
Войти
Т-клетки памяти - это подмножество Т-лимфоцитов, которые могут выполнять некоторые из тех же функций, что и В-клетки памяти. Их происхождение неясно.
Антиген- специфические Т-клетки памяти, специфичные к вирусам или другим микробным молекулам, могут быть обнаружены как в подгруппах Т- CM, так и в T EM. Хотя большая часть информации в настоящее время основана на наблюдениях в подмножестве цитотоксических Т-клеток ( CD8- положительных), похоже, существуют аналогичные популяции как для вспомогательных Т-клеток ( CD4- положительных), так и для цитотоксических Т-клеток. Основная функция клеток памяти - усиление иммунного ответа после реактивации этих клеток путем повторного введения соответствующего патогена в организм. Важно отметить, что эта область интенсивно изучается, и некоторая информация может быть пока недоступна.
Клоны Т-клеток памяти, экспрессирующие специфический Т-клеточный рецептор, могут сохраняться в нашем организме десятилетиями. Поскольку Т-клетки памяти имеют более короткий период полураспада, чем наивные Т-клетки, непрерывная репликация и замена старых клеток, вероятно, вовлечены в процесс поддержания. В настоящее время механизм поддержания Т-клеток памяти до конца не изучен. Активация через рецептор Т-клеток может иметь значение. Обнаружено, что Т-клетки памяти могут иногда реагировать на новые антигены, что может быть вызвано внутренним разнообразием и широтой мишеней связывания рецепторов Т-клеток. Эти Т-клетки могут перекрестно реагировать на антигены окружающей среды или резидентные антигены в нашем организме (например, бактерии в нашем кишечнике) и размножаться. Эти события помогут сохранить популяцию Т-клеток памяти. Механизм перекрестной реактивности может быть важным для Т-клеток памяти в тканях слизистой оболочки, поскольку эти участки имеют более высокую плотность антигена. Для тех, кто находится в крови, костном мозге, лимфоидных тканях и селезенке, более важным может быть передача сигналов гомеостатических цитокинов (включая IL-17 и IL-15 ) или главного комплекса гистосовместимости II (MHCII).
Т-клетки памяти претерпевают разные изменения и играют разные роли на разных этапах жизни человека. При рождении и в раннем детстве Т-клетки периферической крови в основном являются наивными Т-лимфоцитами. В результате частого воздействия антигена происходит накопление популяции Т-клеток памяти. Это этап генерации памяти, который длится от рождения до примерно 20-25 лет, когда наша иммунная система сталкивается с наибольшим количеством новых антигенов. На следующей стадии гомеостаза памяти количество Т-клеток памяти стабилизируется за счет поддержания гомеостаза. На этом этапе иммунный ответ больше смещается в сторону поддержания гомеостаза, поскольку обнаруживается мало новых антигенов. На этом этапе также становится важным наблюдение за опухолью. На более поздних этапах жизни, около 65-70 лет, иммунологическая стадия наступает, в какой стадии иммунного дисрегуляция, наблюдаются снижение функциональных Т - клеток и повышенная восприимчивость к патогенам.
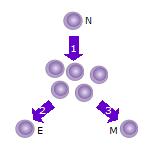 Модель On-Off-On: 1. После того, как наивная Т-клетка (N) встречает антиген, она активируется и начинает пролиферировать ( делиться ) на множество клонов или дочерних клеток. 2. Некоторые из клонов Т-клеток будут дифференцироваться в эффекторные Т-клетки (Е), которые будут выполнять функцию этой клетки (например, продуцировать цитокины в случае хелперных Т-клеток или вызывать уничтожение клеток в случае цитотоксических Т-клеток ). 3. Некоторые из клеток будут формировать Т-клетки памяти (M), которые будут выживать в неактивном состоянии в организме хозяина в течение длительного периода времени, пока они снова не встретятся с тем же антигеном и не активируются.
Модель On-Off-On: 1. После того, как наивная Т-клетка (N) встречает антиген, она активируется и начинает пролиферировать ( делиться ) на множество клонов или дочерних клеток. 2. Некоторые из клонов Т-клеток будут дифференцироваться в эффекторные Т-клетки (Е), которые будут выполнять функцию этой клетки (например, продуцировать цитокины в случае хелперных Т-клеток или вызывать уничтожение клеток в случае цитотоксических Т-клеток ). 3. Некоторые из клеток будут формировать Т-клетки памяти (M), которые будут выживать в неактивном состоянии в организме хозяина в течение длительного периода времени, пока они снова не встретятся с тем же антигеном и не активируются. По состоянию на апрель 2020 года родственные отношения между эффекторными Т-клетками и Т-клетками памяти неясны. Существуют две конкурирующие модели. Одна называется моделью On-Off-On. Когда наивные Т-клетки активируются связыванием Т-клеточного рецептора (TCR) с антигеном и его нижележащим сигнальным путем, они активно пролиферируют и образуют большой клон эффекторных клеток. Эффекторные клетки подвергаются активной секреции цитокинов и другой эффекторной активности. После клиренса антигена некоторые из этих эффекторных клеток образуют Т-клетки памяти либо случайным образом, либо выбираются на основе их превосходной специфичности. Эти клетки вернутся из активной эффекторной роли в состояние, более похожее на наивные Т-клетки, и будут снова «включены» при следующем воздействии антигена. Эта модель предсказывает, что эффекторные Т-клетки могут переходить в Т-клетки памяти и выживать, сохраняя способность к пролиферации. Он также предсказывает, что определенный профиль экспрессии генов будет следовать паттерну включения-выключения во время наивной, эффекторной стадий и стадий памяти. Доказательства, подтверждающие эту модель, включают обнаружение генов, связанных с выживанием и хомингом, которые следуют паттерну экспрессии «включено-выключено», включая альфа-рецептор интерлейкина-7 (IL-7Rα), Bcl-2, CD26L и другие.
 Модель дифференцировки развития: в этой модели Т-клетки памяти генерируют эффекторные Т-клетки, а не наоборот.
Модель дифференцировки развития: в этой модели Т-клетки памяти генерируют эффекторные Т-клетки, а не наоборот. Другая модель - это модель дифференциации развития. Эта модель утверждает, что эффекторные клетки, продуцируемые высокоактивированными наивными Т-клетками, будут подвергаться апоптозу после клиренса антигена. Вместо этого Т-клетки памяти продуцируются наивными Т-клетками, которые активируются, но никогда не входят в полную силу на эффекторную стадию. Потомки Т-клеток памяти не активируются полностью, потому что они не так специфичны к антигену, как расширяющиеся эффекторные Т-клетки. Исследования, посвященные истории деления клеток, показали, что длина теломер и активность теломеразы были уменьшены в эффекторных Т-клетках по сравнению с Т-клетками памяти, что позволяет предположить, что Т-клетки памяти не претерпевали такого большого клеточного деления, как эффекторные Т-клетки, что несовместимо с модель On-Off-On. Повторная или хроническая антигенная стимуляция Т-лимфоцитов, такая как ВИЧ-инфекция, может вызвать усиление эффекторных функций, но ухудшить память. Также было обнаружено, что сильно пролиферирующие Т-клетки с большей вероятностью будут генерировать короткоживущие эффекторные клетки, тогда как минимально пролиферированные Т-клетки будут формировать более долгоживущие клетки.
Эпигенетические модификации участвуют в замене наивных Т-клеток. Например, в Т-клетках памяти CD4 + положительные модификации гистонов маркируют ключевые гены цитокинов, которые активируются во время вторичного иммунного ответа, включая IFNγ, IL4 и IL17A. Некоторые из этих модификаций сохранялись после клиренса антигена, создавая эпигенетическую память, которая позволяет более быструю активацию при повторной встрече с антигеном. Для CD8 + Т-клеток памяти некоторые эффекторные гены, такие как IFNγ, не будут экспрессироваться, но они транскрипционно готовы к быстрой экспрессии при активации. Кроме того, усиление экспрессии некоторых генов также зависит от силы исходной передачи сигналов TCR для потомства Т-клеток памяти, что коррелирует с активацией регуляторного элемента, который напрямую изменяет уровень экспрессии гена.
Исторически считалось, что Т-клетки памяти принадлежат либо к эффекторному, либо к подтипу центральной памяти, каждый со своим собственным отличительным набором маркеров клеточной поверхности (см. Ниже). Впоследствии были обнаружены многочисленные дополнительные популяции Т-клеток памяти, включая резидентные в тканях Т-клетки памяти (Т- RM), стволовые Т- клетки памяти Т- СКМ и Т-клетки виртуальной памяти. Единственная объединяющая тема для всех подтипов Т-клеток памяти заключается в том, что они являются долгоживущими и могут быстро увеличиваться до большого количества эффекторных Т-клеток при повторном воздействии на их родственный антиген. Посредством этого механизма они обеспечивают иммунную систему «памятью» против ранее встреченных патогенов. Т-клетки памяти могут быть CD4 + или CD8 + и обычно экспрессируют CD45RO и в то же время лишены CD45RA.
Было предложено множество других субпопуляций Т-клеток памяти. Исследователи изучили стволовые клетки памяти T SCM. Подобно наивным Т-клеткам, Т- клетки SCM представляют собой CD45RO-, CCR7 +, CD45RA +, CD62L + ( L-селектин ), CD27 +, CD28 + и IL-7Rα +, но они также экспрессируют большие количества CD95, IL-2Rβ, CXCR3 и LFA. -1, и показывают многочисленные функциональные признаки, характерные для ячеек памяти.