
Войти
 Сукцессия после нарушения : бореальный лес один год (слева) и два года (справа) после лесного пожара.
Сукцессия после нарушения : бореальный лес один год (слева) и два года (справа) после лесного пожара.Экологическая сукцессия - это процесс изменения видовой структуры экологическое сообщество с течением времени. Временной масштаб может составлять десятилетия (например, после лесного пожара) или даже миллионы лет после массового вымирания.
Сообщество начинается с относительно небольшого числа растений и животных-первопроходцев и развивается за счет увеличения сложность, пока она не станет стабильной или самовоспроизводящейся как кульминационное сообщество. «Двигатель» преемственности, причина изменения экосистемы - это воздействие устоявшихся организмов на их собственную среду обитания. Следствием жизни является иногда незаметное, а иногда и явное изменение собственного окружения.
Это явление или процесс, посредством которого экологическое сообщество претерпевает более или менее упорядоченные и предсказуемые изменения после нарушение или первоначальное заселение новой среды обитания. Сукцессия может быть инициирована либо образованием новой, незанятой среды обитания, например, потоком лавы или сильным оползнем, либо некоторой формой нарушения сообщества, например, в результате пожара, сильного метания ветра или лесозаготовок. Сукцессия, которая начинается в новых средах обитания, не подверженная влиянию ранее существовавших сообществ, называется первичной сукцессией, тогда как сукцессия, которая следует за разрушением ранее существовавшего сообщества, называется вторичной сукцессией.
Последовательность была одной из первых теории, выдвинутые в экологии. Экологическая преемственность была впервые задокументирована в дюнах Индианы на северо-западе Индианы и остается основой многих экологических наук.
Предшественники идеи экологической преемственности восходят к началу 19 века. Французский естествоиспытатель Адольф Дюро де ла Малль первым применил слово «сукцессия» в отношении развития растительности после вырубки леса. В 1859 г. Генри Дэвид Торо написал обращение под названием «Последовательность лесных деревьев», в котором он описал последовательность в дубово-сосновом лесу. «Наблюдателям давно известно, что белки зарывают орехи в землю, но я не знаю, чтобы кто-нибудь таким образом учел регулярную смену лесов». Австрийский ботаник Антон Кернер опубликовал исследование о последовательности растений в бассейне реки Дунай в 1863 году.
 Дюны Индианы на озере Мичиган, которые стимулировали развитие Коулза его теорий экологической преемственности
Дюны Индианы на озере Мичиган, которые стимулировали развитие Коулза его теорий экологической преемственности Генри Чендлер Коулз в Чикагском университете разработал более формальный понятие преемственности. Вдохновленный исследованиями датских дюн Юджена Уорминга, Коулз изучил развитие растительности на песчаных дюнах на берегах озера Мичиган (Индиана Дюнс ). Он признал, что растительность на дюнах разного возраста можно интерпретировать как разные стадии общей тенденции развития растительности на дюнах (подход к изучению изменения растительности, позже названный замещением пространства-времени, или хронопоследовательностью исследования). Он впервые опубликовал эту работу в качестве статьи в Ботаническом вестнике в 1899 году («Экологические отношения растительности песчаных дюн озера Мичиган»). В этой классической публикации и последующих статьях он сформулировал идею первичной преемственности и понятие sere - повторяющейся последовательности изменений сообщества, специфичных для конкретных условий окружающей среды.
Примерно с 1900 по 1960 год, однако, в понимании преемственности доминировали теории Фредерика Клементса, современника Коулза, который считал, что серии в высшей степени предсказуемы и детерминированы и сходились на климатическом определено стабильное климакс-сообщество независимо от начальных условий. Клементс явно сравнил сукцессионное развитие экологических сообществ с онтогенетическим развитием отдельных организмов, и его модель часто называют псевдоорганической теорией экологии сообществ. Клементс и его последователи разработали сложную таксономию сообществ и последовательных путей.
Генри Глисон предложил противоположную основу еще в 1920-х годах. Модель Глисона была более сложной и гораздо менее детерминированной, чем модель Клемента. Он наиболее существенно отличается от точки зрения Клемента, предполагая гораздо большую роль факторов случайности и отрицая существование согласованных, строго ограниченных типов сообществ. Глисон утверждал, что распределение видов индивидуально реагирует на факторы окружающей среды, и сообщества лучше всего рассматривать как артефакты сопоставления распределений видов. Идеи Глисона, впервые опубликованные в 1926 году, в значительной степени игнорировались до конца 1950-х годов.
Две цитаты иллюстрируют противоположные взгляды Клементса и Глисона. Клементс писал в 1916 году:
Изучение развития растительности обязательно основывается на предположении, что единица или формирование кульминации является органической сущностью. Как организм, образование возникает, растет, созревает и умирает. Более того, каждое кульминационное образование способно воспроизводиться, повторяя с необходимой точностью этапы своего развития.
— Фредерик Клементс, а Глисон в своей статье 1926 года сказал:
Ассоциация - это не организм, вряд ли даже растительная единица, но просто совпадение.
— Генри ГлисонНа самом деле идеи Глисона больше соответствовали первоначальному мышлению Коулза о преемственности. О различении Клементса между первичной преемственностью и вторичной преемственностью, Коулс писал (1911):
Эта классификация не имеет фундаментального значения, поскольку она разделяет такие тесно связанные явления, как эрозия и отложения, и объединяет такие непохожие вещи, как деятельность человека и оседание земли.
— Генри КоулзБолее точные данные - Управляемое тестирование сукцессионных моделей и теории сообщества обычно началось с работ Роберта Уиттакера и в 1950-х и 1960-х годах. С тех пор теория преемственности стала менее монолитной и более сложной. Дж. Коннелл и Р. Слейер предприняли попытку кодификации сукцессионных процессов по механизму. Британские и североамериканские экологи в значительной степени отказались от идеи стабильной кульминационной растительности, а сукцессионные процессы стали рассматриваться как менее детерминированные, играющие важную роль в исторической непредвиденной ситуации и для альтернативных путей в реальном развитии сообществ. Споры продолжаются относительно общей предсказуемости сукцессионной динамики и относительной важности равновесных и неравновесных процессов. Бывший профессор Гарварда Ф.А. Баззаз ввел понятие масштаба в обсуждение, поскольку он считал, что в локальном масштабе или в масштабе небольшой территории процессы являются стохастическими и неоднородными, но, принимая во внимание более крупные региональные районы, нельзя отрицать определенные тенденции.
На траекторию сукцессионных изменений могут влиять условия участка, характер событий, инициирующих сукцессию (возмущения), взаимодействия присутствующих видов и многое другое стохастический такие факторы, как наличие колонистов или семян или погодные условия во время беспокойства. Некоторые из этих факторов способствуют предсказуемости динамики сукцессии; другие добавляют больше вероятностных элементов. Сегодня два важных фактора возмущения - это действия человека и изменение климата.
В целом, в сообществах в ранней сукцессии будут преобладать быстрорастущие, хорошо рассредоточенные виды (оппортунист, беглец или r-selected истории жизни). По мере продолжения сукцессии эти виды будут иметь тенденцию заменяться более конкурентоспособными (k-selected ) видами.
Тенденции в экосистеме и свойствах сообщества последовательно предлагались, но немногие из них были общими. Например, видовое разнообразие почти обязательно увеличивается во время ранней сукцессии по мере появления новых видов, но может снижаться в более поздней сукцессии, поскольку конкуренция устраняет условно-патогенные виды и приводит к доминированию местных более сильных конкурентов. Чистая первичная продуктивность, биомасса и трофические свойства - все они демонстрируют переменные закономерности в последовательности, в зависимости от конкретной системы и участка.
Раньше считалось, что экологическая сукцессия имеет стабильную конечную стадию, называемую кульминацией, иногда называемую «потенциальной растительностью» участка и определяемую, в первую очередь, местным климатом. Современные экологи отказались от этой идеи в пользу неравновесных представлений о динамике экосистем. Большинство естественных экосистем испытывают нарушение со скоростью, которая делает недостижимым "кульминационный момент" сообщества. Изменение климата часто происходит со скоростью и частотой, достаточными для предотвращения наступления кульминационного состояния. Дополнения к доступным пулам видов посредством расширения ареала и интродукции также могут постоянно изменять сообщества.
Развитие некоторых атрибутов экосистемы, таких как свойства почвы и круговорот питательных веществ, оба зависят от свойств сообщества и, в свою очередь, влияют на дальнейшее сукцессионное развитие. Этот процесс обратной связи может происходить только через столетия или тысячелетия. В сочетании со стохастической природой возмущающих событий и других долгосрочных (например, климатических) изменений такая динамика делает сомнительным, применима ли концепция «кульминации» когда-либо или она особенно полезна при рассмотрении фактической растительности.
Сукцессионная динамика, начинающаяся с заселения территории, которая ранее не была занята экологическим сообществом, например, недавно обнаженные скальные или песчаные поверхности, лава потоки, вновь обнаженные ледниковые пазы и т. д. называются первичной сукцессией. Стадии первичной сукцессии включают пионерные микроорганизмы, растения (лишайники и мхи), травянистую стадию, более мелкие кустарники и деревья. Животные начинают возвращаться, когда есть пища для них. Когда это полностью функционирующая экосистема, она достигает кульминационной стадии сообщества.
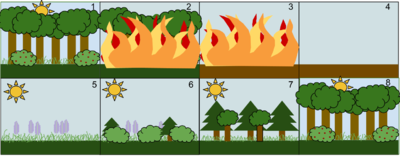 Пример вторичной последовательности по стадиям:. 1. Сообщество стабильного лиственного леса. 2. Возмущение, такое как лесной пожар, уничтожает лес. 3. Огонь сжигает лес дотла. 4. Огонь оставляет после себя пустую, но не уничтоженную землю. 5. Первые отрастают травы и другие травянистые растения. 6. Небольшие кусты и деревья начинают заселять территорию. 7. Быстрорастущие вечнозеленые деревья развиваются в полной мере, тогда как теневыносливые деревья развиваются в подлеске. 8. Недолговечные и непереносимые тени вечнозеленые деревья умирают, когда над ними возвышаются более крупные лиственные деревья. Экосистема теперь вернулась в то же состояние, в котором она началась.
Пример вторичной последовательности по стадиям:. 1. Сообщество стабильного лиственного леса. 2. Возмущение, такое как лесной пожар, уничтожает лес. 3. Огонь сжигает лес дотла. 4. Огонь оставляет после себя пустую, но не уничтоженную землю. 5. Первые отрастают травы и другие травянистые растения. 6. Небольшие кусты и деревья начинают заселять территорию. 7. Быстрорастущие вечнозеленые деревья развиваются в полной мере, тогда как теневыносливые деревья развиваются в подлеске. 8. Недолговечные и непереносимые тени вечнозеленые деревья умирают, когда над ними возвышаются более крупные лиственные деревья. Экосистема теперь вернулась в то же состояние, в котором она началась. Сукцессионная динамика после серьезного нарушения или удаления ранее существовавшего сообщества называется вторичной сукцессией. На динамику вторичной сукцессии сильно влияют условия, предшествующие возмущению, включая развитие почвы, семенные банки, остаточное органическое вещество и остаточные живые организмы. Из-за остаточной плодовитости и ранее существовавших организмов изменение сообщества на ранних стадиях вторичной сукцессии может быть относительно быстрым.
Вторичную сукцессию гораздо чаще наблюдают и изучают, чем первичную. Наиболее распространенные типы вторичной сукцессии включают реакцию на естественные возмущения, такие как пожар, наводнение и сильные ветры, а также на антропогенные нарушения, такие как лесозаготовки и сельское хозяйство. Во вторичной сукцессии почвы и организмы необходимо оставить нетронутыми, чтобы у нового материала был путь к восстановлению.
Например, в фрагментированной старой полевой среде обитания, созданной в восточном Канзасе, древесные растения «колонизировали» более быстро (на единицу площади) на больших и близлежащих участках ".
 Вторичная последовательность: деревья колонизируют невозделываемые поля и луга.
Вторичная последовательность: деревья колонизируют невозделываемые поля и луга. Вторичная последовательность может быстро изменить ландшафт. В 1900-х годах Национальный парк Акадия произошел лесной пожар, уничтоживший большую часть ландшафта. Первоначально в ландшафте росли вечнозеленые деревья. После пожара на этом участке росли кусты не менее года. В конце концов, вместо вечнозеленых растений начали расти лиственные деревья.
Вторичная сукцессия произошла в Национальном парке Шенандоа после наводнения в 1995 г. рек Мурман и Рапидан, уничтожившего растения и животных.
В отличие от вторичной сукцессии, эти типы растительности изменения ионов не зависят от возмущения, а являются периодическими изменениями, возникающими в результате взаимодействия колеблющихся видов или повторяющихся событий. Эти модели модифицируют концепцию кульминации в сторону одного из динамических состояний.
Аутогенная сукцессия может быть вызвана изменениями в почве, вызванными присутствующими там организмами. Эти изменения включают накопление органических веществ в подстилке или гуминовом слое, изменение питательных веществ в почве или изменение pH почвы из-за растущих там растений. Структура самих растений также может изменить сообщество. Например, когда более крупные виды, такие как деревья, созревают, они создают тень на развивающейся лесной подстилке, которая, как правило, исключает светолюбивые виды. Теневыносливые виды будут вторгаться в этот район.
Аллогенная сукцессия вызвана влиянием внешней среды, а не растительностью. Например, изменения почвы из-за эрозии, выщелачивания или отложения ила и глин могут изменить содержание питательных веществ и водные отношения в экосистемах. Животные также играют важную роль в аллогенных изменениях, поскольку они опылители, разносчики семян и травоядные. Они также могут увеличивать содержание питательных веществ в почве в определенных областях или перемещать почву (как это делают термиты, муравьи и кроты), создавая пятна в среде обитания. Это может создать участки регенерации, благоприятствующие определенным видам.
Климатические факторы могут быть очень важными, но в гораздо более длительном масштабе, чем любые другие. Изменения температуры и режима осадков будут способствовать изменениям в сообществах. По мере потепления климата в конце каждого ледникового периода происходили большие сукцессионные изменения. Растительность тундры и голые ледниковые отложения сменились смешанными широколиственными лесами. парниковый эффект, приводящий к повышению температуры, вероятно, принесет глубокие аллогенные изменения в следующем столетии. Геологические и климатические катастрофы, такие как извержения вулканов, землетрясения, лавины, метеоры, наводнения, пожары и сильный ветер, также вызывают аллогенные изменения.
В 1916 году Фредерик Клементс опубликовал описательную теорию сукцессии и выдвинул ее как общую экологическую концепцию. Его теория преемственности оказала сильное влияние на экологическую мысль. Концепцию Клемента принято называть классической экологической теорией. По словам Клементса, сукцессия - это процесс, включающий несколько этапов:
 Сукцессия или серия прудов A: зарождающиеся растения B: отложения C: зарождающиеся растения растут внутрь, отложения накапливаются D: зарождающиеся и наземные растения E: осадки заполняют пруд, наземные растения захватывают F: растут деревья
Сукцессия или серия прудов A: зарождающиеся растения B: отложения C: зарождающиеся растения растут внутрь, отложения накапливаются D: зарождающиеся и наземные растения E: осадки заполняют пруд, наземные растения захватывают F: растут деревья  A hydrosere community
A hydrosere community A seral Сообщество - это промежуточный этап в развитии экосистемы. приближаясь к своему кульминационному сообществу. Во многих случаях до достижения кульминационных состояний развивается более одной стадии развития. prisere - это набор seres, составляющих развитие области от не покрытых растительностью поверхностей до кульминационного сообщества. В зависимости от субстрата и климата встречаются разные серы.
Теория преемственности была разработана в основном ботаниками. Исследование преемственности применительно ко всем экосистемам было начато в трудах Рамона Маргалефа, в то время как публикация Юджина Одума Стратегии развития экосистемы считается его формальной отправная точка.
Жизнь животных также претерпевает изменения с изменением сообществ. В лишайниковой стадии фауна редка. Он состоит из нескольких клещей, муравьев и пауков, обитающих в трещинах и щелях. Качественный рост фауны происходит на стадии разнотравья. Животные, обнаруженные на этом этапе, включают нематод, личинок насекомых, муравьев, пауков, клещей и т. Д. Популяция животных увеличивается и диверсифицируется по мере развития лесного климаксового сообщества. Фауна состоит из беспозвоночных, таких как слизни, улитки, черви, многоножки, многоножки, муравьи, жуки; и позвоночные животные, такие как белки, лисы, мыши, кроты, змеи, различные птицы, саламандры и лягушки.
Последовательность микроорганизмов, включая грибы и бактерии, встречающиеся в микробиологической среде обитания, известная как микросекцессия или серула. Как и в случае с растениями, микробная сукцессия может происходить в новых доступных средах обитания (первичная сукцессия ), таких как поверхности листьев растений, недавно обнаженные каменные поверхности (например, ледниковый тилль) или кишки новорожденных животных, а также в нарушенных сообществах ( вторичная последовательность ), например, растущие на недавно мертвых деревьях или помете животных. Сообщества микробов также могут изменяться из-за продуктов, выделяемых присутствующими бактериями. Изменения pH в среде обитания могут создать идеальные условия для обитания в этом районе нового вида. В некоторых случаях новые виды могут превзойти нынешние по питательным веществам, что приведет к гибели основных видов. Изменения также могут происходить в результате микробной сукцессии с вариациями доступности воды и температуры. Теории макроэкологии только недавно были применены к микробиологии, и еще очень многое предстоит понять в этой развивающейся области. Недавнее исследование микробной сукцессии оценило баланс между стохастическими и детерминированными процессами в бактериальной колонизации солончаков хронопоследовательностью. Результаты этого исследования показывают, что, как и в макросессии, ранняя колонизация (первичная последовательность ) в основном зависит от стохастичности, в то время как вторичная последовательность этих бактериальных сообществ в большей степени зависит от детерминированных факторов.
Согласно классической экологической теории, сукцессия прекращается, когда серия достигает равновесия или устойчивого состояния с физической и биотической средой. За исключением серьезных нарушений, он будет сохраняться бесконечно. Эта конечная точка преемственности называется кульминацией.
Последним или стабильным сообществом в сере является климакс-сообщество или климатическая растительность. Он самовоспроизводящийся и находится в равновесии с физической средой обитания. В климаксовом сообществе нет чистого годового накопления органических веществ. Годовое производство и использование энергии в таком сообществе сбалансировано.
Существуют три школы интерпретаций, объясняющих концепцию климакс:
Теория альтернативных стабильных состояний предполагает, что существует не одна конечная точка, а множество, которые переходят друг в друга в течение экологического времени.

Леса, как экологическая система, подвержены процессу сукцессии видов. Существуют «оппортунистические» или «пионерные» виды, которые производят большое количество семян, разносимых ветром, и поэтому могут колонизировать большие пустые участки. Они способны прорастать и расти под прямыми солнечными лучами. После того, как они произвели a, отсутствие прямого солнечного излучения на почву затрудняет развитие их собственных саженцев. Это возможность для теневыносливых видов закрепиться под защитой первопроходцев. Когда пионеры умирают, их заменяют теневыносливые виды. Эти виды способны расти под пологом, а значит, при отсутствии катастроф останутся. По этой причине тогда говорят, что стенд достиг своего апогея. Когда происходит катастрофа, для пионеров снова открывается возможность, при условии, что они присутствуют или находятся в разумных пределах.
Примером видов-первопроходцев в лесах северо-востока Северной Америки являются Betula papyrifera (Белая береза ) и Prunus serotina (Черная вишня ), которые особенно хорошо растут. приспособлены к использованию больших промежутков в лесных пологах, но не переносят тени и в конечном итоге заменяются другими теневыносливыми видами при отсутствии нарушений, которые создают такие промежутки.
В природе вещи не черно-белые, и есть промежуточные стадии. Поэтому нормально, что между двумя крайностями света и тени существует градиент, и есть виды, которые могут действовать как первопроходцы или терпимые, в зависимости от обстоятельств. Чрезвычайно важно знать толерантность видов, чтобы практиковать эффективное лесоводство.
| На Викискладе есть средства массовой информации, связанные с Экологической преемственностью. |
| В Викиучебнике есть книга по теме: Экология / Преемственность и стабильность сообщества |