
Войти
| DLD | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||
| |||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||
| Псевдонимы | DLD, DLDD, DLDH, E3, GCSL, LAD, PHE3, дигидролипоамиддегидрогеназа, дигидролипоамиддегидрогеназа, OGDC-E3 | ||||||||||||||||||||||||
| Внешние идентификаторы | OMIM: 238331 MGI: 107450 HomoloGene: 84 GeneCard: DLD | ||||||||||||||||||||||||
| |||||||||||||||||||||||||
| Orthologs | |||||||||||||||||||||||||
| Виды | Человек | Мышь | |||||||||||||||||||||||
| Entrez | |||||||||||||||||||||||||
| Ensembl | |||||||||||||||||||||||||
| UniProt | |||||||||||||||||||||||||
| RefSeq (m | |||||||||||||||||||||||||
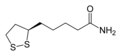
| RefSeq (белок) | Дигидролипоамид дегидрогеназа (DLD), также известная как дигидролипоилдегидрогеназа, митохондриальная, представляет собой фермент , который у человека кодируется геном DLD . DLD - это фермент флавопротеин, который окисляет дигидролипоамид в липоамид. Дигидролипоамиддегидрогеназа (DLD) - митохондриальный фермент, который играет жизненно важную роль в энергетическом обмене у эукариот. Этот фермент необходим для полной реакции по крайней мере пяти различных мультиферментных комплексов. Кроме того, DLD представляет собой флавоэнзим оксидоредуктазу, которая содержит реактивный дисульфидный мостик и кофактор FAD, которые непосредственно участвуют в катализе. Фермент объединяется в прочно связанные гомодимеры, необходимые для его ферментативной активности. Содержание
Структура Белок, кодируемый геном DLD, соединяется с другим белком с образованием димера в центральном метаболическом пути. Несколько аминокислот в каталитическом кармане были идентифицированы как важные для функции DLD, включая R281 и N473. Хотя общая складка человеческого фермента аналогична складке дрожжей, человеческая структура отличается тем, что она имеет две петли, которые простираются от общей белковой структуры в Сайты связывания FAD при связывании с молекулой NAD +, необходимые для катализа, не близки к фрагменту FAD. Однако, когда вместо этого привязан NADH, он укладывается прямо поверх центральной структуры FAD. Существующие структуры hE3 прямо показывают, что вызывающие заболевание мутации возникают в трех местах человеческого фермента: на границе раздела димера, в активном сайте и в FAD. и NAD (+) - сайты связывания. Функция Гомодимер DLD функционирует как компонент E3 пирувата, α-кетоглутарата, и комплексы аминокислоты с разветвленной цепью - дегидрогеназа и система расщепления глицином, все в митохондриальном матриксе. В этих комплексах DLD превращает дигидролипоевую кислоту и NAD + в липоевую кислоту и NADH. DLD также обладает активностью диафоразы, будучи способным катализировать окисление NADH до NAD + с помощью различных акцепторов электронов, таких как O 2, лабильное трехвалентное железо железо, оксид азота и убихинон. Считается, что DLD играет прооксидантную роль, восстанавливая кислород до супероксида или трехвалентного до двухвалентного железа, которое затем катализирует образование гидроксильных радикалов. Активность диафоразы DLD может иметь антиоксидантную роль благодаря своей способности удалять оксид азота и восстанавливать убихинон до убихинола. Известно, что ген дигиролипамиддегидрогеназы имеет несколько вариантов сплайсинга. Подработка Некоторые мутации DLD могут одновременно вызывать потерю основной метаболической активности и усиление подработки протеолитической активности. Подрабатывающая протеолитическая активность DLD проявляется в условиях, которые дестабилизируют гомодимер DLD и снижают его активность DLD. Подкисление митохондриального матрикса в результате ишемии-реперфузионного повреждения может нарушить четвертичную структуру DLD, что приведет к снижению активности дегидрогеназы и повышению активности диафоразы. Подрабатывающая протеолитическая активность DLD может возникать и при патологических условиях. Протеолитическая активность может дополнительно осложнять снижение энергетического метаболизма и увеличение окислительного повреждения в результате снижения активности DLD и увеличения активности диафоразы соответственно. Благодаря своей протеолитической функции DLD удаляет функционально важный домен с N-конца фратаксина, митохондриального белка, участвующего в метаболизме железа и антиоксидантной защите. Клиническое значение Мутации в DLD у людей связаны с тяжелым расстройством детского возраста с задержкой развития, гипотонией и метаболическим ацидозом. Дефицит DLD проявляется в значительной степени вариабельности, что объясняется различным влиянием различных мутаций DLD на стабильность белка и его способность димеризоваться или взаимодействовать с другими компонентами трех комплексов дегидрогеназы α-кетокислоты. Благодаря своей протеолитической функции, DLD вызывает дефицит фратаксина, что приводит к нейродегенеративным и сердечным заболеваниям, атаксии Фридрейха. Дальнейшие исследования надеются оценить, как протеолитическая активность DLD влияет на симптомы дефицита DLD, атаксии Фридрейха и ишемического реперфузионного повреждения, и может ли эта активность быть мишенью для терапии этих состояний. Интерактивная карта путей
Регулирование ферментов Этот белок может использовать морфеин модель аллостерической регуляции. См. Также Ссылки Дополнительная литература Внешние ссылки
Эта статья включает текст из Национальной медицинской библиотеки США, который находится в общественном достоянии. . Последняя правка сделана 2021-05-17 06:05:35
Содержание доступно по лицензии CC BY-SA 3.0 (если не указано иное). | ||||||||||||||||||||||||


 [[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]]
[[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]] [[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]]
[[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]][[]]