
Войти
A сигма-фактор (σ-фактор или фактор специфичности ) - это белок необходимо для инициации транскрипции в бактериях. Это фактор инициации бактериальной транскрипции, который обеспечивает специфическое связывание РНК-полимеразы (RNAP) с промоторами гена . Он гомологичен фактору транскрипции B архей и фактору эукариот TFIIB. Конкретный сигма-фактор, используемый для инициации транскрипции данного гена, будет варьироваться в зависимости от гена и от сигналов окружающей среды, необходимых для инициации транскрипции этого гена. Выбор промоторов РНК-полимеразой зависит от связанного с ним сигма-фактора. Они также обнаруживаются в растительных хлоропластах как часть бактериоподобной пластид-кодируемой полимеразы (PEP).
Сигма-фактор вместе с РНК-полимеразой известен как РНК-полимераза холоэнзим. Каждая молекула холофермента РНК-полимеразы содержит ровно одну субъединицу сигма-фактора, которая в модельной бактерии Escherichia coli является одной из перечисленных ниже. Количество сигма-факторов варьируется между видами бактерий. E. coli имеет семь сигма-факторов. Сигма-факторы различаются по характерным молекулярным массам. Например, σ - это сигма-фактор с молекулярной массой 70 кДа.
Сигма-фактор в холоферментном комплексе РНК-полимеразы необходим для инициации транскрипции, хотя по завершении этой стадии он отделяется от комплекс, и RNAP продолжает удлинение самостоятельно.
В разных условиях окружающей среды используются разные сигма-факторы. Эти специализированные сигма-факторы связывают промоторы генов, соответствующие условиям окружающей среды, увеличивая транскрипцию этих генов.
Сигма-факторы в E. coli :
Существуют также антисигма-факторы, которые подавляют функцию сигма-факторов, и анти-антисигма-факторы, восстанавливающие функция сигма-фактора.
 Организация домена, распознавание промотора и структурная организация семейства σ. (a ) Доменная организация σ-факторов из групп 1, 3 и 4 проиллюстрирована выше σ консенсусной промоторной ДНК E. coli. (b ) Организация E. coli σ70 в комплексе инициации транскрипции РНК-полимеразы. (PDB 4YLN).
Организация домена, распознавание промотора и структурная организация семейства σ. (a ) Доменная организация σ-факторов из групп 1, 3 и 4 проиллюстрирована выше σ консенсусной промоторной ДНК E. coli. (b ) Организация E. coli σ70 в комплексе инициации транскрипции РНК-полимеразы. (PDB 4YLN). По сходству последовательностей большинство сигма-факторов являются σ-подобными (InterPro : IPR000943 ). У них есть четыре основных региона (домена), которые обычно консервативны:
N-конец --------------------- C-конец 1.1 2 3 4
Далее регионы подразделяются. Например, область 2 включает 1,2 и 2,1–2,4.
Домен 1.1 обнаруживается только в «первичных сигма-факторах» (RpoD, RpoS в E.coli; «Группа 1»). Он участвует в обеспечении того, чтобы сигма-фактор связывался с промотором только тогда, когда он находится в комплексе с РНК-полимеразой. Каждый из доменов 2-4 взаимодействует со специфическими элементами промотора и с RNAP. Область 2.4 распознает и связывается с элементом промотора -10 (называемым «блоком Прибнова »). Область 4.2 распознает и связывается с элементом промотора -35.
Не каждый сигма-фактор семейства σ содержит все домены. Группа 2, которая включает RpoS, очень похожа на группу 1, но не имеет домена 1. Группа 3 также не имеет домена 1 и включает σ. В группе 4, также известной как группа с экстрацитоплазматической функцией (ECF), отсутствуют как σ1.1, так и σ3. RpoE является членом.
Другие известные сигма-факторы относятся к типу σ / RpoN (InterPro : IPR000394 ). Они являются функциональными сигма-факторами, но у них существенно различаются первичные аминокислотные последовательности.
|
|
|
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
РНК-полимераза ядра (состоящая из 2 альфа (α), 1 бета (β), 1 бета-прим. β ') и 1 субъединица омега (ω)) связывает сигма-фактор с образованием комплекса, называемого РНК-полимеразой холоэнзимом. Ранее считалось, что холофермент РНК-полимеразы инициирует транскрипцию, тогда как основная РНК-полимераза сама по себе синтезирует РНК. Таким образом, общепринятая точка зрения заключалась в том, что сигма-фактор д. Диссоциировать при переходе от инициации транскрипции к удлинению транскрипции (этот переход называется «уход от промотора»). Эта точка зрения была основана на анализе очищенных комплексов РНК-полимеразы, остановившихся при инициации и при удлинении. Наконец, структурные модели комплексов РНК-полимеразы предсказывают, что по мере того, как растущий продукт РНК становится длиннее, чем ~ 15 нуклеотидов, сигма должна быть «вытеснена» из голофермента, поскольку существует стерическое противоречие между РНК и сигма-доменом. Однако недавнее исследование показало, что σ может оставаться прикрепленным в комплексе с основной РНК-полимеразой, по крайней мере, во время раннего удлинения. В самом деле, феномен паузы проксимального промотора указывает на то, что сигма играет роль во время ранней элонгации. Все исследования согласуются с предположением, что ускользание промотора сокращает время жизни взаимодействия сигма-ядро с очень долгого при инициировании (слишком долгого для измерения в типичном биохимическом эксперименте) до более короткого, измеримого времени жизни при переходе к удлинению.
Долгое время считалось, что сигма-фактор обязательно покидает основной фермент, как только он инициировал транскрипцию, позволяя ему связываться с другим основным ферментом и инициировать транскрипцию в другом сайте. Таким образом, сигма-фактор циклически переходит от одного ядра к другому. Однако Ричард Эбрайт и его сотрудники, используя резонансный перенос энергии флуоресценции, позже показали, что сигма-фактор не обязательно покидает ядро. Вместо этого он меняет свою связь с ядром во время инициации и удлинения. Следовательно, сигма-фактор циклически изменяется между сильно связанным состоянием во время инициирования и слабосвязанным состоянием во время удлинения.
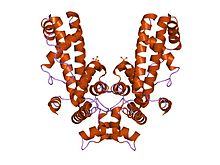 Кристаллическая структура фрагмента сигма-субъединицы РНК-полимеразы Thermus aquaticus, содержащего области от 1.2 до 3.1
Кристаллическая структура фрагмента сигма-субъединицы РНК-полимеразы Thermus aquaticus, содержащего области от 1.2 до 3.1 Кристаллическая структура фрагмента субъединицы сигма70 из РНК-полимеразы Escherichia coli
Кристаллическая структура фрагмента субъединицы сигма70 из РНК-полимеразы Escherichia coli Структура решения sigma70 региона 3 из Thermotoga maritima
Структура решения sigma70 региона 3 из Thermotoga maritima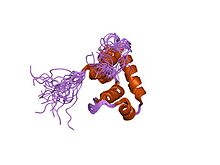 Структура решения sigma70 региона 4 из Thermotoga maritima
Структура решения sigma70 региона 4 из Thermotoga maritima Кристаллическая структура области 4 Escherichia coli sigma70, связанной с ее -35-элементной ДНК
Кристаллическая структура области 4 Escherichia coli sigma70, связанной с ее -35-элементной ДНК