
Войти
| Центр цвета | |
|---|---|
 Область цветного зрения, обозначенная как V8 на верхнем изображении Область цветного зрения, обозначенная как V8 на верхнем изображении | |
| Анатомическая терминология [редактировать в Викиданных ] |
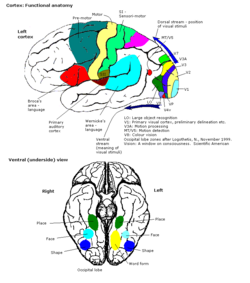
центр цвета - это область в головном мозге, в первую очередь отвечающая за зрительное восприятие и кортикальную обработку цветовых сигналов, принимаемых глазом, что в конечном итоге приводит к цветовому зрению. Считается, что центр окраски у людей расположен в вентральной затылочной доле как часть зрительной системы в дополнение к другим областям, отвечающим за распознавание и обработку определенных зрительных стимулов., например лица, слова и предметы. Многие исследования функциональной магнитно-резонансной томографии (фМРТ) как на людях, так и на макаках обезьянах показали, что цветовые стимулы активируют несколько областей мозга, включая веретенообразную извилину и язычная извилина. Эти области, а также другие, определенные как имеющие роль в обработке цветового зрения, вместе обозначаются областью зрения 4 (V4). Точные механизмы, расположение и функции V4 все еще исследуются.
Первичная часть зрительной коры (V1) расположена в calcarine sulcus, и является первой кортикальной областью, участвующей в визуальной обработке. Он получает визуальный сигнал от латерального коленчатого ядра, расположенного в таламусе. V1 отправляет визуальную информацию, полученную от LGN, в другие области экстрастриарной коры для обработки более высокого порядка. Эта обработка более высокого порядка включает в себя распознавание форм, движения и цвета.
V1 имеет несколько областей, чувствительных к цвету, что указывает на то, что обработка цвета не ограничивается одной областью. Согласно статье доктора Роберта Шепли, V1 играет важную роль в восприятии цвета. Результаты экспериментов с фМРТ показали, что V1 имеет два типа цветочувствительных нейронов: клетки с одним оппонентом и клетки с двумя оппонентами. Эти ячейки являются неотъемлемой частью оппонентского процесса интерпретации цветовых сигналов. Нейроны-одиночки реагируют на большие цветные области. Это удобно для распознавания больших цветных сцен и атмосфер. Для сравнения, клетки двойного противника реагируют на паттерны, текстуры и цветовые границы. Это более важно для восприятия цвета предметов и картинок. Клетки с двойным оппонентом восприимчивы к противоположным входным сигналам от разных колбочек в сетчатке. Это идеально подходит для определения контрастных цветов, например красного и зеленого. [1] Двойные оппонентные клетки особенно важны при вычислении соотношений локальных колбочек на основе визуальной информации из их рецептивных полей.
Цветочувствительные нейроны одиночных оппонентов можно разделить на две категории в зависимости от сигналов, которые они получают от колбочек. : Нейроны LM и нейроны S / (L + M). Три типа колбочек, маленькие (S), средние (M) и длинные (L), обнаруживают разные длины волн в видимом спектре. Клетки S-колбочек могут видеть цвета с короткой длиной волны, что соответствует фиолетовому и синему. Точно так же M-клетки обнаруживают цвета со средней длиной волны, такие как зеленый и желтый, а L-клетки обнаруживают цвета с длинной длиной волны, такие как красный. L-M нейроны, также называемые красно-зелеными оппонентными клетками, получают входные данные от длинноволновых конусов, а не средневолновые. S / (L + M) нейроны получают входные данные от S-клеток, и им противопоставляется сумма входов L- и M-клеток. S / (L + M) нейроны также называют сине-желтыми оппонентными клетками. Противопоставление цветов позволяет визуальной системе интерпретировать различия в цвете, что в конечном итоге более эффективно, чем обработка цветов по отдельности.
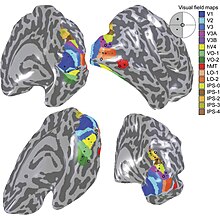 Карта поля зрения первичной зрительной коры и множества Экстрастриальные области.
Карта поля зрения первичной зрительной коры и множества Экстрастриальные области. Первичная зрительная кора V1 отправляет визуальную информацию в экстрастриальные области коры головного мозга для визуальной обработки более высокого порядка. Эти экстрастриарные области коры расположены кпереди от затылочной доли. Основные обозначены как визуальные области V2, V3, V4 и V5 / MT. Каждая область может иметь несколько функций. Недавние открытия показали, что центр окраски не изолирован и не связан с какой-то одной областью зрительной коры. Скорее, есть несколько областей, которые могут играть разные роли в способности обрабатывать цветовые стимулы.
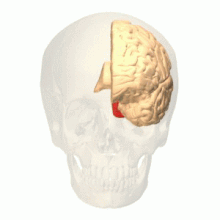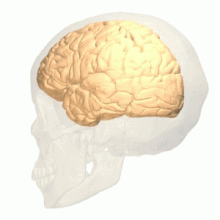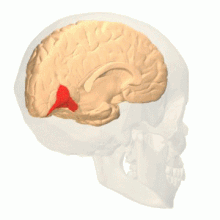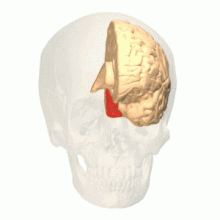 Лингвальная извилина - это гипотетическое расположение V4 у макак. У человека эта область называется hV4.
Лингвальная извилина - это гипотетическое расположение V4 у макак. У человека эта область называется hV4.  Веретенообразная извилина - это гипотетическое расположение V4α, вторичной области для обработки цвета.
Веретенообразная извилина - это гипотетическое расположение V4α, вторичной области для обработки цвета. Анатомические и физиологические исследования установили, что центр окраски начинается в V1 и посылает сигналы экстристрам. области V2 и V4 для дальнейшей обработки. V4 представляет особый интерес из-за силы цветовых рецептивных полей в нейронах. V4 был первоначально идентифицирован в экспериментах со зрительной корой головного мозга макак. Первоначально предполагалось, что в V4 выборочная обработка цвета. Однако позже эта гипотеза была отвергнута в пользу другой гипотезы, которая предполагала, что V4 и другие области вокруг V4 работают вместе, чтобы обрабатывать цвет в виде нескольких областей выборки цвета. После идентификации V4 как цветоселективной области у макак, ученые начали поиск гомологичной структуры в коре головного мозга человека. Используя томографию головного мозга с помощью фМРТ, ученые обнаружили три основные области, стимулируемые цветом: V1, область в вентральной затылочной доле, в частности язычная извилина, которая была обозначена как V4 человека или hV4, и еще одна область, расположенная спереди в веретенообразной извилине, обозначенная как как V4α.
Назначение V4 динамически менялось по мере проведения новых исследований. Поскольку V4 сильно реагирует на цвет как у макак, так и у людей, он стал предметом интереса ученых. Первоначально область V4 была связана с избирательностью цвета, но новые данные показали, что V4, как и другие области зрительной коры, восприимчивы к различным входным сигналам. Нейроны V4 восприимчивы к ряду свойств, таких как цвет, яркость и текстура. Он также участвует в обработке формы, ориентации, кривизны, движения и глубины.
Фактическая организация hV4 в коре все еще исследуется. У макаки V4 охватывает спинную и вентральную затылочную долю. Эксперименты на людях показали, что V4 охватывает только вентральную часть. Это привело к отличию hV4 от V4 макаки. Недавнее исследование Winawer et al. Анализ измерений фМРТ для картирования hV4 и вентральной затылочной области показал, что различия между субъектами, использованными для картирования hV4, сначала были связаны с ошибкой инструментария, но Винавер утверждал, что синусы в головном мозге мешают измерениям fMRI. Были протестированы две модели для hV4: одна модель имела hV4 полностью на вентральной стороне, а вторая модель имела hV4, разделенную на дорсальный и вентральный отделы. Был сделан вывод, что картировать активность hV4 по-прежнему сложно и что необходимы дальнейшие исследования. Однако другие свидетельства, такие как поражения вентральной затылочной доли, вызывающие ахроматопсию, предполагают, что вентральная затылочная область играет важную роль в цветовом зрении.
поиск человеческого эквивалента V4 привел к открытию других областей, которые стимулировались цветом. Наиболее значимой была передняя область вентральной затылочной доли, впоследствии названная V4α. Дальнейшие эксперименты с фМРТ показали, что V4α имел другую функцию, чем V4, но работал с ним совместно. V4α участвует в ряде процессов и активен во время задач, требующих упорядочения цветов, изображений, знаний о цвете, цветовых иллюзий и цвета объекта.
Области V4 и V4α - отдельные объекты, но из-за их непосредственной близости в веретенообразной извилине эти две области часто вместе называют комплексом V4. Исследование комплекса V4 обнаружило, что различные хроматические стимуляции активируют либо область V4, либо область V4α, а некоторые параметры стимуляции активируют оба. Например, изображения с естественной окраской активировали V4α сильнее, чем V4. Неестественно окрашенные изображения активировали как V4α, так и V4 одинаково. Был сделан вывод, что два подразделения взаимодействуют друг с другом для создания цветных изображений, но они также функционально разделены.
Исследование Nunn et al. об активации V4-комплекса у людей с зрительной синестезией при прослушивании произнесенных слов использовали для прогнозирования местоположения цветового центра. Синестезия - это явление, при котором сенсорный раздражитель вызывает автоматическую и непроизвольную реакцию в виде другого ощущения. В этом исследовании изучали людей, которые видят цвета, слыша слова, чтобы увидеть, можно ли отследить цветовую реакцию в определенной области коры головного мозга. Результаты фМРТ показали, что левая веретенообразная извилина, область, соответствующая V4, активировалась, когда испытуемые говорили. Они также обнаружили одновременную активацию V4α. В областях V1 и V2 активность была незначительной. Эти результаты подтвердили существование V4-комплекса у людей как области, специализирующейся на цветовом зрении.
V2, также называемый prestriate cortex, как полагают, играет небольшую роль при обработке цвета путем проецирования сигналов из V1 в V4-комплекс. Присутствуют ли цветоселективные клетки в V2, все еще исследуется. Некоторые исследования с оптической визуализацией обнаружили небольшие кластеры клеток с селективным красно-зеленым цветом в V1 и V2, но не сине-желтых клеток. Другие исследования показали, что V2 активируется цветовыми стимулами, но не цветом после изображений. [8] V4 также имеет обратную связь с V2, предполагая, что существует определенная сеть связи между несколькими областями зрительной коры. Когда ГАМК, ингибирующий нейромедиатор, вводили в клетки V4, клетки V2 испытывали значительное снижение возбудимости.
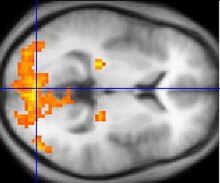 фМРТ, проявляющие активность в первичная зрительная кора V1.
фМРТ, проявляющие активность в первичная зрительная кора V1. Функциональная магнитно-резонансная томография, или для краткости фМРТ, была ключевой в определении цветоселективных областей в зрительной коре. ФМРТ может отслеживать активность мозга, измеряя кровоток по всему мозгу. Области, к которым притекает больше крови, указывают на возникновение нейрональной активности. Это изменение кровотока называется гемодинамической реакцией. Среди преимуществ фМРТ - динамическое картирование корковых процессов в реальном времени. Однако фМРТ не может отслеживать фактическое срабатывание нейронов, которое происходит в миллисекундной шкале времени, но она может отслеживать гемодинамическую реакцию, которая происходит в секундах. Этот метод идеален для отслеживания нейронов, избирательных по цвету, поскольку восприятие цвета приводит к визуальному остаточному изображению, которое можно наблюдать в нейронах, которое длится около 15 секунд.
Sakai et al. использовали фМРТ, чтобы наблюдать, коррелирует ли активация веретенообразной извилины с восприятием цвета и последующим изображением. Субъекты в исследовании Сакаи были помещены в аппарат фМРТ и впоследствии подверглись воздействию различных зрительных стимулов. Субъектам была показана серия из трех изображений, в то время как фМРТ использовалась для сосредоточения внимания на гемодинамике веретенообразной извилины. Первое изображение представляло собой узор из шести цветных кругов. Следующие два изображения были ахроматическими. На одном из изображений был серый крест, а на другом изображении были те же шесть кругов, что и на первом изображении, за исключением шести оттенков серого, которые коррелировали с цветными изображениями. Испытуемые переключались между кругом и крестиком. Во время перекрестных изображений субъект воспринимал остаточное изображение. Результаты эксперимента показали, что при просмотре цветного изображения субъектом наблюдалось значительное увеличение активности веретенообразной извилины. Это дало больше доказательств существования центра окраски за пределами первичной зрительной коры.
Церебральная ахроматопсия - хроническое заболевание, при котором человек не может видеть цвет, но он все еще может распознавать форму и форму. Церебральная ахроматопсия отличается от врожденной ахроматопсии тем, что она вызвана повреждением коры головного мозга, а не аномалиями клеток сетчатки. Поиски центра окраски были мотивированы открытием того, что поражения вентральной затылочной доли приводят к дальтонизу, а также идеей о том, что в коре головного мозга существуют определенные области. Многие исследования показали, что поражения в областях, обычно определяемых как центры окраски, таких как V1, V2 и V4-комплекс, приводят к ахроматопсии. Церебральная ахроматопсия возникает после повреждения язычной или веретенообразной извилины, областей, связанных с hV4. Эти травмы включают физическую травму, инсульт и рост опухоли. Одна из основных инициатив по обнаружению цветового центра в зрительной коре - выявление причины и возможное лечение церебральной ахроматопсии.
 Моделирование церебральной ахроматопсии.
Моделирование церебральной ахроматопсии. Степень симптомов и повреждений у разных людей разная. Если у человека полная ахроматопсия, то все его поле зрения лишено цвета. Человек с дисхроматопсией или неполной ахроматопсией имеет симптомы, аналогичные полной ахроматопсии, но в меньшей степени. Это может произойти у людей, у которых была ахроматопсия, но мозг восстановился после травмы, частично восстановив цветовое зрение. Человек может видеть определенные цвета. Однако во многих случаях восстановления нет. Наконец, человек с гемахроматопсией видит половину своего поля зрения в цвете, а другую половину - в сером. Зрительное полуполе, контралатеральное поражению язычной или веретенообразной извилины, выглядит серым, а ипсилатеральное зрительное полуполе появляется в цвете. Различия в симптомах подчеркивают необходимость понимания архитектуры цветового центра, чтобы лучше диагностировать и возможное лечение церебральной ахромотопсии.
| Викискладе есть медиафайлы, относящиеся к визуальной области V4. |