
Войти
 Цвета оппонента на основе эксперимента. Дейтеранопы видят небольшую разницу между двумя цветами в центральном столбце.
Цвета оппонента на основе эксперимента. Дейтеранопы видят небольшую разницу между двумя цветами в центральном столбце. 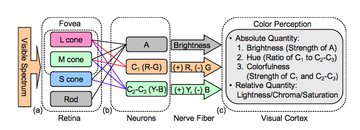 Диаграмма процесса оппонента
Диаграмма процесса оппонента 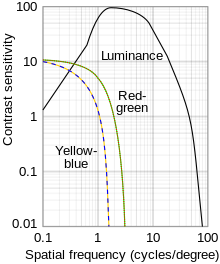 Логарифмический график функций пространственной контрастной чувствительности для яркости и хроматического контраста
Логарифмический график функций пространственной контрастной чувствительности для яркости и хроматического контраста процесс-оппонент - это теория цвета, которая утверждает, что человеческая зрительная система интерпретирует информацию о цвете путем обработки сигналов от колбочек и стержневые клетки антагонистически. Существует некоторое перекрытие длин волн из света, на которое реагируют три типа колбочек (L для длинноволнового, M для средневолнового и S для коротковолнового света)., поэтому для зрительной системы более эффективно записывать различия между ответами колбочек, а не индивидуальный ответ каждого типа колбочек. Теория цвета оппонента предполагает, что существует три канала оппонента, фоторецепторы конуса связаны вместе, образуя три противоположные цветовые пары: красный против зеленый, синий по сравнению с желтым и черным по сравнению с белым (последний тип - ахроматический и обнаруживает изменение света и темноты, или яркость ). Впервые он был предложен в 1892 году немецким физиологом Эвальдом Герингом.
Когда люди слишком долго смотрят на яркий цвет, например красный, и отводят взгляд на белое поле, они воспринимают зеленый цвет. Активация одного члена пары подавляет активность другого. Эта теория также помогает объяснить некоторые типы нарушения цветового зрения. Например, люди с дефектами дихроматического должны сопоставить тестовое поле, используя только два основных цвета. В зависимости от недостатка они будут путать красный и зеленый или синий и желтый. Теория процесса оппонента объясняет цветовое зрение как результат нейронной связи фоторецепторов. Теория процесса оппонента применима к разным уровням нервной системы. Как только нейронная система выходит за пределы сетчатки в мозг, природа клетки изменяется, и клетка реагирует на нее противоположным образом. Например, зеленый и красный фоторецепторы могут отправлять сигнал в сине-красную клетку противника дальше вместе с системой. Ответы на один цвет канала оппонента антагонистичны откликам на другой цвет. То есть противоположные цвета оппонента никогда не воспринимаются вместе - нет «зеленовато-красного» или «желтовато-синего».
В то время как трихроматическая теория определяет способ, которым сетчатка глаза позволяет зрительной системе определять цвет с помощью трех типов колбочек, теория процесса оппонента учитывает механизмы которые получают и обрабатывают информацию от конусов. Хотя первоначально считалось, что теории трехцветных и оппонентных процессов расходятся, позже стало понятно, что механизмы, ответственные за оппонентный процесс, получают сигналы от трех типов колбочек и обрабатывают их на более сложном уровне.
Помимо колбочек, которые обнаруживают свет, попадающий в глаз, биологическая основа теории оппонента включает два других типа клеток: биполярные клетки и ганглиозные клетки. Информация от колбочек передается биполярным клеткам сетчатки, которые могут быть клетками в противоборствующем процессе, которые преобразовывают информацию от колбочек. Затем информация передается ганглиозным клеткам, из которых существует два основных класса: магноцеллюлярные или большие клеточные слои и парвоцеллюлярные или мелкоклеточные слои. слои. Парвоцеллюлярные клетки, или P-клетки, обрабатывают большую часть информации о цвете и делятся на две группы: одна обрабатывает информацию о различиях между возбуждением L и M колбочек, а вторая обрабатывает различия между S колбочками и комбинированный сигнал от обоих L и M конусов. Первый подтип клеток отвечает за обработку красно-зеленых различий, а второй - за сине-желтые различия. P-ячейки также передают информацию об интенсивности света (сколько его имеется) благодаря своим рецептивным полям.
Иоганн Вольфганг фон Гете впервые изучил физиологический эффект противоположных цветов в своей Теории цветов в 1810 году. Гете расположил свое цветовое колесо симметрично, «ибо цвета, диаметрально противоположные друг другу на этой диаграмме, - это те, которые взаимно вызывают друг друга в глазах. Таким образом, желтый требует пурпурного, оранжевого - синего, красного, зеленого и. наоборот: таким образом, снова все промежуточные градации взаимно вызывают друг друга ".
Эвальд Геринг предложил оппонентскую теорию цвета в 1892 году. Он думал, что красный, желтый, зеленый и синий цвета являются особенными в том смысле, что любой другой цвет может можно описать как их смесь, и что они существуют в противоположных парах. То есть воспринимается либо красный, либо зеленый, и никогда не зеленовато-красный: хотя желтый - это смесь красного и зеленого в теории цвета RGB, глаз не воспринимает его как таковой. В 1957 году Лео Гурвич и Доротея Джеймсон предоставили количественные данные для теории цветного оппонента Геринга. Их метод был назван подавлением оттенка. Эксперименты по отмене оттенка начинаются с цвета (например, желтого) и пытаются определить, сколько цвета противника (например, синего) одного из компонентов исходного цвета необходимо добавить, чтобы исключить любой намек на этот компонент из исходного цвета. В 1959 году Светичин и МакНикол записали данные с сетчатки рыб и сообщили о трех различных типах клеток: один реагировал гиперполяризацией на все световые стимулы независимо от длины волны и был назван клеткой светимости. Вторая клетка ответила гиперполяризацией на коротких волнах и деполяризацией на средних и длинных волнах. Это было названо ячейкой цветности. Третья ячейка, также ячейка цветности, ответила гиперполяризацией на довольно коротких длинах волн, достигая пика около 490 нм, и деполяризацией на длинах волн более примерно 610 нм. Светичин и МакНикол назвали ячейки цветности желто-синими и красно-зелеными оппонентными цветными ячейками. Сходные хроматически или спектрально противоположные клетки, часто включающие пространственную противоположность (например, красный «в центре» и зеленый «вне» окружения), были обнаружены в сетчатке позвоночных и латеральном коленчатом ядре (LGN) в течение 1950-х годов и 1960-е годы Де Валуа и др., Визеля и Хьюбеля и др. После лидерства Светичина клетки стали широко называть цветными клетками оппонента, красно-зелеными и желто-синими. В течение следующих трех десятилетий спектрально противоположные клетки продолжали обнаруживаться в сетчатке приматов и LGN. В литературе используются различные термины для описания этих ячеек, включая хроматически противоположные или -оппонентные, спектрально-противоположные или -оппонентные, цвет оппонента, цвет оппонента, реакцию оппонента и просто оппонент.
Теория цвета оппонента может быть применена к компьютерному зрению и реализована как и.
Другие применили идею противодействия стимуляции помимо зрительных систем, описанную в статья по теории процесса оппонента. В 1967 г. расширил эту концепцию, чтобы отразить широкий спектр противоборствующих процессов в биологических системах. В 1970 году Соломон и Корбит расширили общую неврологическую модель процесса оппонента Хурвича и Джеймсона, чтобы объяснить эмоции, наркоманию и мотивацию к работе.
Существует множество споров о том, является ли теория обработки оппонентов лучшим способом объяснить цветовое зрение. Было проведено несколько экспериментов, связанных со стабилизацией изображения (когда вы испытываете потерю границ), которые дали результаты, которые предполагают, что участники видели «невозможные» цвета или цветовые комбинации, которые мы не должны видеть в рамках теории обработки оппонентов; Однако многие критикуют, что это может быть просто иллюзорный опыт. Вместо этого критики и исследователи начали объяснять цветовое зрение ссылками на механизмы сетчатки, а не на обработку оппонентов, которая происходит в зрительной коре головного мозга.
По мере накопления записей от одной клетки многим физиологам и психофизикам стало ясно, что противоположные цвета не могут удовлетворительно объяснить спектрально противоположные ответы отдельных клеток. Например, Джеймсон и Д’Андрейд проанализировали теорию цветов оппонента и обнаружили, что уникальные оттенки не соответствуют спектрально противоположным ответам. Сам Де Валуа резюмировал это: «Хотя нас, как и других, больше всего впечатлило обнаружение ячеек оппонента, в соответствии с предположениями Геринга, когда Zeitgeist в то время категорически противился этому понятию, самые ранние записи выявили несоответствие между Герингом. - Каналы восприятия оппонентов Гурвича-Джеймсона и характеристики реакции оппонентных клеток в латеральном коленчатом ядре макака ». Вальберг вспоминает, что «среди нейрофизиологов стало обычным употребление терминов, связанных с цветом, когда речь идет о клетках-оппонентах, как в обозначениях« красные клетки »,« зеленые клетки выключены »... В ходе дискуссии... некоторые психофизики были счастливы чтобы увидеть то, что они считали оппонентом, подтвержденным на объективном, физиологическом уровне. Следовательно, не было никаких сомнений в том, чтобы связать уникальные и полярные цветовые пары непосредственно с противоположностью колбочек. Несмотря на доказательства обратного... в учебниках до сих пор повторяется неправильное представление о связи восприятия уникального оттенка непосредственно с периферическими процессами оппонента колбочек. Аналогия с гипотезой Геринга была продвинута еще дальше и подразумевала, что каждый цвет в оппонентной паре уникальных цветов может быть идентифицирован либо с возбуждением, либо с подавлением одного и того же типа оппонентной клетки ». Webster et al. и Wuerger et al. окончательно подтвердили, что спектрально противоположные ответы отдельных ячеек не совпадают с цветами оппонентов уникального оттенка.
В 2013 году Придмор утверждал, что большинство красно-зеленых ячеек, описанных в литературе, на самом деле кодируют красно-голубые цвета. Таким образом, клетки кодируют дополнительные цвета вместо цветов-оппонентов. Придмор сообщил также о зелено-пурпурных клетках сетчатки и V1. Таким образом, он утверждал, что красно-зеленые и сине-желтые ячейки следует вместо этого называть «зелено-пурпурными», «красно-голубыми» и «сине-желтыми» дополнительными ячейками. Пример дополнительного процесса можно увидеть, глядя на красный (или зеленый) квадрат в течение сорока секунд, а затем сразу же глядя на белый лист бумаги. Затем наблюдатель видит голубой (или пурпурный) квадрат на чистом листе. Этот дополнительный цвет остаточное изображение легче объяснить теорией трехцветного цвета, чем традиционной теорией цвета RYB; в теории процесса оппонента усталость путей, способствующих развитию красного цвета, создает иллюзию голубого квадрата.