
Войти
| TEAD1 | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
| Идентификаторы | ||||||||||||||||||||||||||||||
| Псевдонимы | TEAD1, AA, NTEF-1, REF1, TCF-13, TCF13, TEAD-1, TEF-1, фактор транскрипции домена TEA 1 | |||||||||||||||||||||||||||||
| Внешние идентификаторы | OMIM: 189967 MGI: 101876 HomoloGene: 2418 GeneCards: TEAD1 | |||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
| Orthologs | ||||||||||||||||||||||||||||||
| Species | Human | Mouse | ||||||||||||||||||||||||||||
| Entrez | ||||||||||||||||||||||||||||||
| Ensembl | ||||||||||||||||||||||||||||||
| UniProt | ||||||||||||||||||||||||||||||
| RefSeq (мРНК) |
Фактор усилителя транскрипции TEF-1, также известный как член семейства 1 домена ТЕА ( TEAD1 ) и фактор транскрипции 13 (TCF-13 ) - это белок, который у человека кодируется геном TEAD1 . TEAD1 был первым идентифицированным членом семейства транскрипционных факторов семейства TEAD. 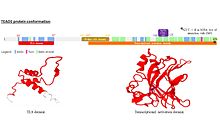 Содержание
Структура Все члены семейства TEAD разделяют высококонсервативное связывание с ДНК домен называется доменом TEA. Этот ДНК-связывающий домен имеет консенсусную последовательность ДНК 5’-CATTCCA / T-3 ’, которая называется элементом MCAT. Была идентифицирована трехмерная структура TEA-домена [5]. Его конформация близка к гомеодомену и содержит 3 α-спирали (H1, H2 и H3). Именно спираль H3 позволяет белкам TEAD связывать ДНК. Другой консервативный домен TEAD1 расположен на С-конце белка. Он позволяет связывать кофакторы и был назван связывающим доменом YAP1, потому что именно его способность связывать этот хорошо известный кофактор белков TEAD привела к его идентификации. Действительно, белки TEAD не могут сами по себе вызывать экспрессию генов. Они должны связываться с кофакторами, чтобы иметь возможность действовать. Распределение в тканях TEAD1 экспрессируется в различных тканях, включая скелетные мышцы, поджелудочную железу, плаценту, легкие и сердце. Ортологи Белки TEAD обнаруживаются во многих организмах под разными названиями, принимая на себя разные функции. Например, у Saccharomyces cerevisiae TEC-1 регулирует мобильный элемент TY1 и участвует в росте псевдогифа (удлиненная форма, которую дрожжи принимают при выращивании в условиях бедности питательными веществами). У Aspergillus nidulans белок домена ТЕА ABAA регулирует дифференцировку конидиофоров. У дрозофилы фактор транскрипции Scalloped участвует в развитии крыльевого диска, выживании и росте клеток. Наконец, на Xenopus было продемонстрировано, что ортолог TEAD1 регулирует дифференцировку мышц. Функция
Посттранскрипционные модификации Протеинкиназа A (pKA) может фосфорилировать TEAD1 по серину 102 после домена TEA. Это фосфорилирование необходимо для активации транскрипции гена α MyHC. Протеинкиназа C (pKC) фосфорилирует TEAD1 по серину и треонину рядом с последней альфа-петлей в TEA-домене. Это фосфорилирование снижает связывание TEAD1 с энхансером GTIIC. TEAD1 может быть пальмитоилирован консервативным цистеином на С-конце белка. Эта посттрансляционная модификация имеет решающее значение для правильного сворачивания белков TEAD и их стабильности. Кофакторы Белки TEAD требуют кофакторов для индукции транскрипции генов-мишеней. TEAD1 взаимодействует со всеми членами семейства SRC коактиваторов стероидных рецепторов. В клетках HeLa TEAD1 и SRC индуцируют экспрессию генов, TEAD1 взаимодействует с PARP (поли-АДФ-рибозная полимераза), чтобы регулировать экспрессию α-актина в гладких мышцах. PARP может также АДФ-рибозилировать белки TEAD и делать контекст хроматина благоприятным для транскрипции посредством модификации гистонов, SRF (фактор ответа сыворотки) и TEAD1 вместе регулируют экспрессию гена. белки TEAD и MEF2 (фактор усиления миоцитов 2) взаимодействовать физически. Связывание MEF2 с ДНК индуцирует и усиливает рекрутирование TEAD1 в последовательностях MCAT, которые примыкают к сайтам связывания MEF2. Это привлечение приводит к репрессии промотора MLC2v (легкая цепь миозина 2 v) и βMHC (тяжелая цепь β-миозина). TEAD1 и фосфопротеин MAX взаимодействуют in vivo и in vitro. Как только этот комплекс образуется, эти два белка могут регулировать экспрессию гена тяжелой цепи альфа-миозина (α-MHC). Четыре белка, подобные остаткам (VGLL), способны взаимодействовать со всеми TEAD. Точная функция взаимодействия TEAD и VGLL все еще плохо изучена. Было показано, что комплексы TEAD / VGLL1 способствуют независимой от якорения пролиферации клеток в клеточных линиях рака простаты, предполагая роль в прогрессировании рака. Более того, взаимодействие VGLL2 с TEAD1 активирует мышечный промотор при дифференцировке C2C12 и усиливает MyoD-опосредованную миогенность в 10T1 / 2. Наконец, комплекс TEAD / VGLL4 действует как репрессор транскрипции по умолчанию. Взаимодействие между YAP (Yes Associated Protein 65), TAZ, паралогом коактиватора транскрипции к YAP, и всеми белками TEAD было продемонстрировано как in vitro, так и in vitro. естественным образом. В обоих случаях взаимодействие белков приводит к увеличению транскрипционной активности TEAD. YAP / TAZ являются эффекторами пути опухолевого супрессора Hippo, который ограничивает рост органов, сдерживая пролиферацию клеток и способствуя апоптозу у млекопитающих, а также у дрозофилы. Роль в раке Анализ транскриптома рака базы данных (www.ebi.ac.uk/gxa) показали, что TEAD1 не регулируется при нескольких типах рака. Во-первых, при саркоме Капоши уровень TEAD1 увеличивается в 300 раз. Более того, повышение экспрессии TEAD может быть обнаружено при базальном раке молочной железы, карциноме маточной трубы и опухолях половых клеток. В противном случае при других типах рака экспрессия TEAD снижается, например, при других типах рака груди, а также при раке почек или мочевого пузыря. Эта двойная роль может быть объяснена различными мишенями и дифференциальной регуляцией генов-мишеней факторами транскрипции TEAD. Наконец, недавние исследования показали, что TEAD1 и YAP при раке яичников могут вызывать стволовые клетки и химиорезистентность. и этот генетический вариант белка TEAD и YAP обогащен некоторыми видами рака. Ссылки Дополнительная литература
Последняя правка сделана 2021-06-09 05:44:12
Содержание доступно по лицензии CC BY-SA 3.0 (если не указано иное). | |||||||||||||||||||||||||||||



