
Войти
 Трехдоменное древо жизни, показывающее разделение доменов бактерий, архей и эукариот. См. Статью « Микроорганизмы» для получения дополнительных сведений.
Трехдоменное древо жизни, показывающее разделение доменов бактерий, архей и эукариот. См. Статью « Микроорганизмы» для получения дополнительных сведений. Ученые, пытающиеся реконструировать эволюционную историю, столкнулись с проблемой того факта, что гены иногда могут передаваться между удаленными ветвями на древе жизни. Это движение генов может происходить посредством горизонтального переноса генов (ГПГ), шифруя информацию, на которую биологи полагались при реконструкции филогении организмов. И наоборот, HGT также может помочь ученым восстановить древо жизни и датировать его. Действительно, перенос генов можно использовать как филогенетический маркер или как доказательство современности организмов-доноров и организмов-реципиентов, а также как след исчезнувшего биоразнообразия.
ГПГ случается очень редко - на уровне отдельного организма такое событие крайне маловероятно. Однако в более широком масштабе эволюционной истории эти события происходят с некоторой регулярностью. С одной стороны, это вынуждает биологов отказаться от использования отдельных генов в качестве хороших маркеров истории жизни. С другой стороны, это почти неизведанный большой источник информации о прошлом.
Три основных ранних ветви древа жизни интенсивно изучались микробиологами, потому что первые организмы были микроорганизмами. Микробиологи (во главе с Карлом Вёзе ) ввели термин домен для трех основных ветвей этого дерева, где домен - это филогенетический термин, сходный по значению с биологическим царством. Для воссоздания этого древа жизни оказалась полезной последовательность гена, кодирующая небольшую субъединицу рибосомной РНК (SSU рРНК, 16s рРНК ), и дерево (как показано на рисунке) в значительной степени полагается на информацию от этого единственного гена.
Эти три области жизни представляют собой основные эволюционные линии ранней клеточной жизни и в настоящее время включают бактерии, археи (одноклеточные организмы, внешне похожие на бактерии) и эукарии. Эукария включает только организмы с четко определенным ядром, такие как грибы, простейшие, и все организмы в царствах растений и животных (см. Рисунок).
Геном, наиболее часто используемым для построения филогенетических взаимоотношений у микроорганизмов, является ген малой субъединицы рибосомной РНК, поскольку его последовательности, как правило, сохраняются среди членов с близкими филогенетическими расстояниями, но при этом достаточно изменчивы, чтобы можно было измерить различия. РРНК SSU как мера эволюционных расстояний была впервые предложена Карлом Вёзе при формулировании первого современного «древа жизни», и его результаты привели его к тому, что он предложил архей в качестве третьей области жизни. Однако недавно было высказано мнение, что гены SSU рРНК также могут переноситься горизонтально. Хотя это может быть редкостью, такая возможность требует тщательного изучения валидности филогенетических деревьев, основанных на рРНК SSU.
Недавние открытия «безудержного» ГПГ у микроорганизмов и обнаружение горизонтального движения даже генов небольшой субъединицы рибосомной РНК заставили биологов усомниться в точности по крайней мере ранних ветвей в дереве и даже поставить под сомнение обоснованность деревья как полезные модели того, как происходит ранняя эволюция. Фактически считается, что ранняя эволюция произошла от сообщества предшественников, способных обмениваться большими молекулами, когда ГПГ был стандартом. Этот латеральный перенос гена произошел также за дарвиновским порогом, после того, как была установлена наследственность или вертикальный перенос гена. «Сравнение последовательностей предполагает недавний горизонтальный перенос многих генов между различными видами, в том числе через границы филогенетических « доменов ». Таким образом, определение филогенетической истории вида не может быть сделано окончательно путем определения эволюционных деревьев для отдельных генов». Таким образом, HGT является потенциальным вмешивающимся фактором при выводе филогенетических деревьев из последовательности одного гена. Например, если две отдаленно родственные бактерии обменялись генами, филогенетическое древо, включающее эти виды, покажет, что они тесно связаны, даже если большинство других генов существенно разошлись. По этой причине важно использовать другую информацию для вывода о филогенезе, такую как наличие или отсутствие генов, или, что более часто, включать как можно более широкий спектр генов для анализа.
В своей статье « Выкорчевывая Древо Жизни» У. Форд Дулиттл обсуждает Последнего Универсального Общего Предка - корень Древа Жизни - и проблемы с этой концепцией, поставленные HGT. Он описывает микроорганизм Archaeoglobus fulgidus как аномалию по отношению к филогенетическому дереву, основанному на коде фермента HMGCoA-редуктазы - организм определенно архей со всеми липидами клетки и аппаратом транскрипции, ожидаемым от архея, но его гены HMGCoA являются бактериального происхождения. В статье Дулиттл говорит, что, хотя сейчас широко признано, что митохондрии у эукариот происходят из альфа-протеобактериальных клеток, а хлоропласты - из проглоченных цианобактерий,
«... больше не безопасно предполагать, что это были единственные латеральные переносы генов, которые произошли после появления первых эукариот. Только у более поздних многоклеточных эукариот мы знаем об определенных ограничениях на горизонтальный обмен генами, таких как появление разделенных ( и защищенные) половые клетки. ..
Если бы не происходил какой-либо боковой перенос генов, все эти отдельные генные деревья имели бы одинаковую топологию (один и тот же порядок ветвления), и все предковые гены в корне каждого дерева присутствовали бы в последнем универсальном общем предке, одиночная древняя клетка. Но обширный перенос означает, что это не так: деревья генов будут различаться (хотя многие из них будут иметь области схожей топологии), и никогда не было бы ни одной клетки, которую можно было бы назвать последним универсальным общим предком... "
Дулиттл предположил, что универсальный общий предок не мог быть одним конкретным организмом, а должен был быть рыхлым, разнообразным конгломератом примитивных клеток, которые развивались вместе. Эти ранние клетки, каждая с относительно небольшим количеством генов, во многом различались и свободно меняли местами свои гены. В конце концов, из этих эклектических клеток произошли три области жизни, какими мы их знаем сегодня: бактерии, археи и эукариоты. Эти домены теперь узнаваемо различны, потому что большая часть передачи генов, которая все еще происходит, происходит внутри этих доменов, а не между ними. Биолог Питер Гогартен усилил эти аргументы и предположил, что метафора дерева не соответствует данным недавнего исследования генома, и что биологи должны вместо этого использовать «метафору мозаики для описания различных историй, объединенных в отдельных геномах, и использовать [ ] метафора сети для визуализации богатого обмена и совместных эффектов ГПГ среди микробов ».
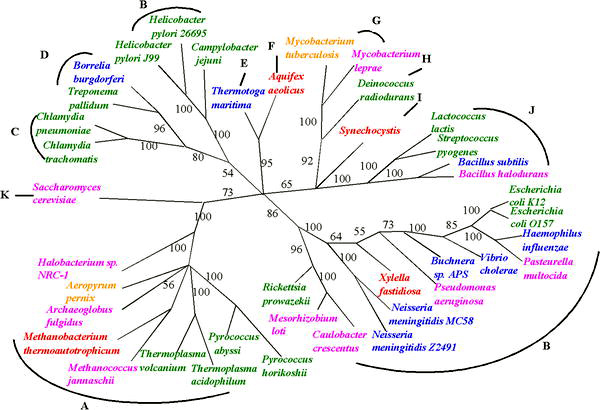 Дерево на основе алгоритма медианного дерева. Ветви с начальной (статистической) оценкой менее 50% были свернуты. Три области жизни: (A) археи, (B – J) бактерии и (K) эукариоты. Виды помечены разными цветами в зависимости от предполагаемых показателей HGT: красный,gt; 4%; желтый, 3–4%; розовый, 2–3%; синий, 1–2%; зеленый, lt;1%. Таксономические ярлыки: (A) Euryarchaea, (B) Proteobacteria, (C) Chlamydiae, (D) Spirochaetes, (E) Thermotogae, (F) Aquificae, (G) Actinobacteria, (H) Deinococcus, (I) Cyanobacteria, (J)) Firmicutes и (K) Fungi.
Дерево на основе алгоритма медианного дерева. Ветви с начальной (статистической) оценкой менее 50% были свернуты. Три области жизни: (A) археи, (B – J) бактерии и (K) эукариоты. Виды помечены разными цветами в зависимости от предполагаемых показателей HGT: красный,gt; 4%; желтый, 3–4%; розовый, 2–3%; синий, 1–2%; зеленый, lt;1%. Таксономические ярлыки: (A) Euryarchaea, (B) Proteobacteria, (C) Chlamydiae, (D) Spirochaetes, (E) Thermotogae, (F) Aquificae, (G) Actinobacteria, (H) Deinococcus, (I) Cyanobacteria, (J)) Firmicutes и (K) Fungi. Несмотря на неопределенность в восстановлении филогенеза до истоков жизни, прогресс в восстановлении древа жизни наблюдается перед лицом неопределенностей, порожденных ГПГ. Неопределенность любого предполагаемого филогенетического дерева, основанного на единственном гене, может быть решена с помощью нескольких общих генов или даже данных из целых геномов. Один из таких подходов, иногда называемый «мультилокусным типированием», был использован для вывода филогенных деревьев для организмов, которые обмениваются генами, таких как бактерии менингита.
Джонатан Эйзен и Клэр Фрейзер отметили, что:
"При построении древа жизни анализ полных геномов начал дополнять, а в некоторых случаях улучшать исследования, ранее проводившиеся с одним или несколькими генами. Например, недавние исследования полных бактериальных геномов показали, что гипертермофильные виды не имеют глубокого разветвления; если это правда, это ставит под сомнение идею о том, что первые формы жизни были термофилами. Анализ генома эукариотического паразита Encephalitozoon cuniculi подтверждает предположение, что группа Microsporidia не является протистами с глубоким ветвлением, а на самом деле является представители грибного царства. Анализ генома может даже помочь установить взаимосвязи внутри видов, например, путем предоставления новых генетических маркеров для популяционных генетических исследований бактерий, вызывающих сибирскую язву или туберкулез. Во всех этих исследованиях это дополнительные данные, предоставляемые полным геномом последовательность, которая позволяет отделить филогенетический сигнал от шума. Это не значит, что древо жизни теперь разрешено - мы на Я опробовал небольшое количество геномов, и многие группы еще не затронуты »
Эти подходы позволяют оценить относительную частоту ГПГ; относительно низкие значения, которые наблюдались, предполагают, что «дерево» по-прежнему является допустимой метафорой для эволюции - но дерево украшено «паутиной» горизонтально перенесенных генов. Это главный вывод исследования более 40 полных геномных последовательностей микробов в 2005 году, проведенного Фан Ге, Ли-Сан Ван и Чунхён Ким. По их оценкам, частота событий HGT составляет около 2% основных генов на геном. Подобные полногеномные подходы к оценке эволюции также способствуют прогрессу в выявлении очень ранних событий на древе жизни, например, предположение, что эукариоты возникли путем слияния двух полных, но очень разных геномов прокариот: одного из бактерий и одного из архейной клетки..
Такая гипотеза слияния организмов о происхождении сложных ядерных клеток была выдвинута Линн Маргулис с использованием совершенно иных рассуждений о симбиозе между бактерией и археном, возникающим в древнем консорциуме микробов.
В то время как HGT часто рассматривается как вызов при реконструкции древа жизни, альтернативная точка зрения состоит в том, что напротив он предоставляет дополнительную ценную информацию для его реконструкции.
Во-первых, для организма-реципиента HGT представляет собой мутацию ДНК, как и другие, и поэтому его можно смоделировать и использовать для реконструкции и укоренения деревьев.
Во-вторых, необходимо, чтобы реципиент, полученный с помощью HGT, жил одновременно или в более позднее время с донором. Как следствие, есть информация о сроках диверсификации в HGT. Это тем более примечательно, что основной обычный источник для датировки в живом мире, летопись окаменелостей, отсутствует именно там, где ГПГ в изобилии, в микробном мире.
В-третьих, он предоставляет информацию о исчезнувшем биоразнообразии, потому что перенос, вероятно, от вымерших видов.