
Войти
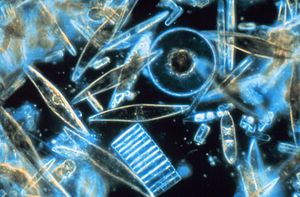 Диатомовые водоросли способны синтезировать кварцевое стекло in vivo.
Диатомовые водоросли способны синтезировать кварцевое стекло in vivo.Биогенный кремнезем (bSi), также упоминается как опал, биогенный опал или аморфный опалин кремнезем, образует один из наиболее распространенных биогенных минералов. Например, микроскопические частицы кремнезема, называемые фитолитами, можно найти в травах и других растениях. Кремнезем представляет собой аморфный оксид металла , образованный сложными неорганическими процессами полимеризации. Этим он отличается от других основных биогенных минералов, содержащих карбонат и фосфат, которые встречаются в природе в виде кристаллических ионо-ковалентных твердых веществ (например, солей ), осаждение продиктовано равновесием растворимости. Химически bSi представляет собой гидратированный диоксид кремния (SiO 2 · nH 2 O), который важен для многих растений и животных.
Силикат или кремниевая кислота (H4SiO 4), является важным питательным веществом в океане. В отличие от других основных питательных веществ, таких как фосфат, нитрат или аммоний, которые необходимы почти всему морскому планктону, силикаты являются основные химические требования для очень специфической биоты, включая диатомеи, радиолярии, силикофлагелляты и кремнистые губки. Эти организмы извлекают растворенный силикат из поверхностных вод открытого океана для накопления в них твердого кремнезема (SiO 2) или опаловых структур скелета (т. Е. Твердых частей биоты). Некоторые из наиболее распространенных кремнистых структур, наблюдаемых на поверхности клеток организмов, выделяющих кремнезем, включают: спикулы, чешуйки, твердые пластинки, гранулы, створки и другие сложные геометрические формы, в зависимости от рассматриваемых видов
. Можно выделить источники растворенного кремнезема в морскую среду:
После гибели организма часть кремнеземистого скелетного материала растворяется, поскольку он оседает через водная толща, обогащающая глубинные воды растворенным кремнеземом. Некоторые из кремнистых чешуек могут также сохраняться с течением времени в виде микрофоссилий в глубоководных отложениях, открывая окно в современный и древний планктон / протистов сообществ. Этот биологический процесс действовал, по крайней мере, с раннего палеозоя времени, чтобы регулировать баланс кремнезема в океане: радиолярии (кембрий / ордовик - Голоцен ), диатомеи (мел - голоцен ) и силикофлагеллаты (мел - голоцен ) образуют основные составляющие океана в глобальный биогенный цикл кремнезема на протяжении геологического времени. На диатомовые водоросли приходится 43% первичной продукции океана, и они ответственны за большую часть добычи кремнезема из океанских вод в современном океане на протяжении большей части последних пятидесяти миллионов лет. Напротив, океаны юрского и более раннего возраста были охарактеризованы радиоляриями как основные типы, использующие кремнезем. В настоящее время радиолярии являются вторыми (после диатомовых) основными продуцентами взвешенного аморфного кремнезема в водах океана. Их распространение колеблется от Арктики до Антарктики, причем наиболее многочисленны они в экваториальной зоне. Например, в экваториальных водах Тихого океана можно наблюдать около 16 000 особей на кубический метр.
Цикличность силикатов в последнее десятилетие привлекает все большее внимание ученых по следующим причинам. Во-первых, широко распространено мнение, что в современном морском круговороте кремнезема преобладают диатомеи для фиксации и экспорта твердых частиц (включая органический углерод ) из эвфотической зоны в глубокий океан с помощью процесса, известного как биологический насос. В результате диатомеи и другие организмы, выделяющие кремнезем, играют решающую роль в глобальном углеродном цикле и обладают способностью влиять на концентрации в атмосфере CO2в различных временных масштабах, посредством секвестрации CO2в океане. Эта связь между биогенным кремнеземом и органическим углеродом вместе со значительно более высоким потенциалом сохранения биогенных кремнистых соединений по сравнению с органическим углеродом делает записи о накоплении опала очень интересными для палеоокеанографии и палеоклиматологии. Во-вторых, накопление биогенного кремнезема на морском дне содержит много информации о том, где в океане экспортное производство происходило в масштабах времени от сотен до миллионов лет. По этой причине записи отложений опалов предоставляют ценную информацию о крупномасштабных океанографических реорганизациях в геологическом прошлом, а также о палеопродуктивности. И, наконец, среднее время пребывания в океане силиката составляет примерно 10 000–15 000 лет. Такое относительно короткое время пребывания делает концентрации и потоки силикатов в океане чувствительными к ледниковым / межледниковым возмущениям и, таким образом, отличным показателем для оценки изменений климата.
Остатки диатомовых водорослей и других организмов, потребляющих кремний, обнаружены в виде опаловых отложений в пелагических глубоководных отложениях. Пелагические отложения, содержащие значительные количества кремнеземистых биогенных остатков, обычно называют кремнистыми илами. Кремнистый ил особенно распространен в современном океане в высоких широтах северного и южного полушарий. Отличительной особенностью распределения кремнистого ила является ок. Полоса шириной 200 км, протянувшаяся через Южный океан. Некоторые экваториальные районы апвеллинга, где питательные вещества обильны, а продуктивность высока, также характеризуются местными кремнистыми илами. Кремнистые илы состоят в основном из остатков диатомовых водорослей и радиолярий, но могут также включать другие кремнистые организмы, такие как силикофлагеллаты и губчатые спикулы. Ил диатомовых водорослей встречается в основном в высокоширотных районах и вдоль некоторых окраин континентов, тогда как илы радиолярий более характерны для экваториальных районов. Кремнистый ил модифицируется и превращается во время захоронения в слоистые кремни.
Диатомовые водоросли как в пресной, так и в соленой воде извлекают кремнезем из воды для использования в качестве компонента своих клеточных стенок. Аналогичным образом, некоторые голопланктонные простейшие (Radiolaria ), некоторые губки и некоторые растения (лист фитолиты ) используют кремний как конструкционный материал. Кремний, как известно, необходим цыплятам и крысам для роста и развития скелета. Кремний находится в соединительных тканях, костях, зубах, коже, глазах, железах и органы.
BSi представляет собой кремнезем, который получают из растворенного кремнезема. BSi может накапливаться «непосредственно» в морских отложениях (через экспорт) или переноситься обратно в растворенный кремнезем в толще воды.
Все чаще изотопные отношения кислорода (O: O) и кремния (Si: Si) анализируются из BSi, сохраненного в озерных и морских отложениях, для получения записей о прошлых изменениях климата. и круговорот питательных веществ (Де Ла Роша, 2006; Ленг и Баркер, 2006). Это особенно ценный подход, учитывая роль диатомовых в глобальном круговороте углерода. Кроме того, изотопный анализ BSi полезен для отслеживания прошлых изменений климата в таких регионах, как Южный океан, где сохранилось мало биогенных карбонатов.
Реки и подводные гидротермальные эманации поставляют в морскую среду 6,1 × 10 г SiO 2 лет. Примерно две трети этого поступления кремнезема хранится в отложениях континентальной окраины и глубоководных. Кремнистые глубоководные отложения, расположенные ниже Антарктической конвергенции (зона конвергенции ), содержат около 25% кремнезема, поступающего в океаны (т.е. 1,6 × 10 г SiO 2 год) и, следовательно, образуют один из основных поглотителей кремнезема на Земле. Самые высокие темпы накопления биогенного кремнезема в этом районе наблюдаются в Южной Атлантике, и за последние 18000 лет они достигли 53 см. Кир. Кроме того, обширное накопление биогенного кремнезема было зарегистрировано в глубоководных отложениях Берингова моря, Охотского моря и Субарктического северной части Тихого океана. Общая скорость накопления биогенного кремнезема в этих регионах составляет около 0,6 × 10 г SiO 2 год, что эквивалентно 10% растворенного кремнезема, поступающего в океаны. Континентальные окраинные районы апвеллинга, такие как Калифорнийский залив, побережье Перу и Чили, характеризуются одними из самых высоких темпов накопления биогенного кремнезема в мире. Например, для Калифорнийского залива сообщалось о скорости накопления биогенного кремнезема 69 г SiO 2 2 28 / см / тыс. Лет. Из-за ограниченного по горизонтали характера этих зон быстрого биогенного накопления кремнезема, на районы апвеллинга приходится только около 5% растворенного кремнезема, поступающего в океаны. Наконец, чрезвычайно низкие темпы накопления биогенного кремнезема наблюдались в обширных глубоководных отложениях Атлантического, Индийского и Тихого океанов, что сделало эти океаны несущественными для глобального морского баланса кремнезема.
Крупномасштабная океаническая циркуляция оказывает прямое влияние на осаждение опалов. Циркуляция в Тихом океане (характеризующемся бедными питательными веществами поверхностными водами и глубокими водами, богатыми питательными веществами) и Атлантического океана способствует производству / сохранению кремнезема и карбоната соответственно. Например, отношения Si / N и Si / P увеличиваются от Атлантики к Тихому и Южному океану, отдавая предпочтение производителям опала по сравнению с карбонатом. Следовательно, современная конфигурация крупномасштабной океанической циркуляции привела к локализации основных зон захоронения опалов в экваториальной части Тихого океана, в системах апвеллинга на восточной границе и, безусловно, в самой важной - Южном океане.
В водах современного Тихого и Южного океанов обычно наблюдается увеличение отношения Si / N на средней глубине, что приводит к увеличению экспорта опала (~ увеличению производства опала). В Южном океане и северной части Тихого океана эта взаимосвязь между экспортом опала и отношением Si / N переключается с линейной на экспоненциальную для отношений Si / N более 2. Это постепенное увеличение важности силиката (Si) по сравнению с азотом (N) имеет огромные последствия для биологического производства океана. Изменение соотношения питательных веществ способствует выбору диатомей в качестве основных продуцентов по сравнению с другими (например, кальцифицирующими) организмами. Например, эксперименты с микрокосмом продемонстрировали, что диатомовые водоросли являются суперконкурентами DSi и доминируют над другими продуцентами с концентрацией DSi выше 2 мкМ. Следовательно, предпочтение будет отдаваться экспорту опала по сравнению с экспортом карбоната, что приведет к увеличению производства опала. Южный океан и северная часть Тихого океана также демонстрируют максимальные отношения потоков биогенный силикат / C органический, и, таким образом, состоят в обогащении биогенным силикатом по сравнению с экспортным потоком C органических. Это совокупное увеличение сохранности и экспорта опалов делает Южный океан наиболее важным поглотителем DSi на сегодняшний день.
В современных водах Тихого и Южного океанов промежуточные и глубокие воды характеризуются более высоким содержанием DSi по сравнению с Атлантический океан. Эта межбассейновая разница в DSi увеличивает потенциал сохранения опалов в Тихом и Южном океанах по сравнению с их атлантическими аналогами. Истощенные воды атлантического DSi имеют тенденцию производить относительно менее окремненных организмов, что оказывает сильное влияние на сохранение их панцирей. Этот механизм лучше всего проиллюстрирован при сравнении систем апвеллинга в Перу и северо-западной Африке. Отношение растворение / продуктивность в атлантическом апвеллинге намного выше, чем в тихоокеанском апвеллинге. Это связано с тем, что прибрежные воды источника апвеллинга намного богаче DSi у побережья Перу, чем у северо-западной Африки.
Отложения Южного океана являются основным поглотителем биогенного кремнезема (50-75% общего объема океана, содержащего 4,5 × 10 г SiO 2 год; DeMaster, 1981), но лишь незначительным поглотителем органического углерода (<1% of the oceanic 2 × 10 g of organic C yr). These relatively high rates of biogenic silica accumulation in the Southern Ocean sediments (predominantly beneath the Polar Front) relative to organic carbon (60:1 on a weight basis) results from the preferential preservation of biogenic silica in the Antarctic water column. In contrast to what was previously thought, these high rates of biogenic silica accumulation are not the result from high rates of первичная продукция. Биологическая продукция в Южном океане сильно ограничена из-за низких уровней освещенности в сочетании с глубокими смешанными слоями и / или ограниченными количествами питательных микроэлементов, например, железо. Эта предпочтительная сохранность биогенного диоксида кремния по сравнению с органическим углеродом проявляется в неуклонно увеличивающемся соотношении диоксид кремния / органический углерод в зависимости от глубины в толще воды. Около тридцати пяти процентов биогенный кремнезем, образующийся в зоне эвфотической, выживает при растворении в поверхностном слое; в то время как только 4% органического углерода улетучивается микробное разложение в этих приповерхностных водах. Следовательно, во время осаждения через толщу воды происходит значительное разделение органического углерода и кремнезема. Накопление биогенного кремнезема на морском дне составляет 12% от продукции на поверхности, в то время как скорость накопления органического углерода на морском дне составляет исключительно <0.5% of the surface production. As a result, polar sediments account for most of the ocean's biogenic silica accumulation, but only a small amount of the sedimentary organic-carbon flux.
 Кизельгур при ярком поле освещении на световом микроскопе . Кизельгур состоит из диатома клеточных стенок, примера биогенного кремнезема. Кремнезем синтезируется в клетке диатомовых водорослей путем полимеризации кремниевой кислоты. Это изображение частиц диатомовой земли в воде имеет масштаб 6,236 пикселей / мкм, все изображение покрывает область размером примерно 1,13 на 0,69 мм.
Кизельгур при ярком поле освещении на световом микроскопе . Кизельгур состоит из диатома клеточных стенок, примера биогенного кремнезема. Кремнезем синтезируется в клетке диатомовых водорослей путем полимеризации кремниевой кислоты. Это изображение частиц диатомовой земли в воде имеет масштаб 6,236 пикселей / мкм, все изображение покрывает область размером примерно 1,13 на 0,69 мм. Среднесуточная скорость BSi сильно зависит от региона :
Аналогично, интегрированная годовая добыча BSi сильно зависит от региона:
Производство BSi контролируется:
Растворение BSi контролируется:
Сохранность BSi измеряется:
Сохранение BSi контролируется:
В кратере Гусева на Марсе Mars Exploration Rover Spirit случайно обнаружил опаловый кремнезем. Одно из его колес ранее было обездвижено и, таким образом, эффективно закапывало марсианский реголит, когда оно тащилось за марсоходом. Более поздний анализ показал, что кремнезем свидетельствует о гидротермальных условиях.