
Войти
| Сучианцы Временной диапазон: ранний триас - недавний,249,7–0 млн лет PreꞒ Ꞓ О S D C п Т J K Стр. N | |
|---|---|
 | |
| Paratypothorax andressorum, ветрозавр | |
 | |
| Prestosuchus chiniquensis, ранний лорикатан | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Класс: | Рептилии |
| Клэйд: | Псевдозухии |
| Клэйд: | Сучия Кребс, 1974 |
| Подгруппы | |
| |
Suchia является клады из археозавров, содержащих большинство pseudosuchians ( крокодилов и их вымерших родственников). Несбитт (2011) определил ее как наименее инклюзивную кладу, содержащую Aetosaurus ferratus, Rauisuchus tiradentes, Prestosuchus chiniquensis и Crocodylus niloticus (живой нильский крокодил). Как правило, единственная группа псевдозухий, которая не указана в Сушиа, - это семейство Ornithosuchidae, хотя по крайней мере один анализ классифицирует орнитосухид как близких родственников эрпетосухид (которые обычно считаются таковыми) и этозавров (которые являются таковыми по определению группы). Фитозавры также исключены из Сучии, хотя неясно, можно ли их вообще квалифицировать как псевдозухий.
Существуют некоторые разногласия по поводу того, какие черты, если таковые имеются, можно использовать, чтобы отличить сучков от архозавров, не относящихся к такому же типу. Анатомические особенности, которые развиваются в основе группы и, таким образом, могут использоваться для характеристики группы, известны как синапоморфии. В двух наиболее всеобъемлющих недавних исследованиях взаимоотношений архозавров, Nesbitt (2011) и Ezcurra (2016), пришли к разным выводам о том, какие синапоморфии характеризуют Сучию. Тем не менее, они согласились с одним признаком, который, вероятно, можно было бы квалифицировать как синапоморфию сучии: пяточный бугорок, который был значительно шире, чем его высота.
Поскольку Сучия является кладой, она определяется отношениями, а не общими характеристиками между ее членами. Тем не менее, некоторые черты встречаются у большинства сучианов, но не у орнитосухид, фитозавров или других архозавров и родственников архозавров, поэтому их можно использовать, чтобы определить, является ли рептилия сициантом или нет. Эти черты распространены по всему телу, но разные палеонтологи расходятся во мнениях относительно того, были ли они истинными синапоморфиями (производными отличительными чертами).
В то время как Бентон и Кларк (1988) обнаружили многочисленные характеристики клады, позже переименованной в Сучию, в исследовании Пола Серено 1991 года о взаимоотношениях архозавров утверждалось, что многие из них также присутствовали у орнитосухид, фитозавров или авеметатарсалий, поэтому их нельзя рассматривать как признаки, которые диагностировать Сучию (по его определению группы). Вместо этого Серено перечислил одну синапоморфию для Сучии. Заглазничный - чешуйчатый бар, который отделяет верхнюю и нижнюю временную fenestrae в задней части черепа, является коротким. В результате нижнее височное отверстие приобретает треугольную форму. Однако Серено также отметил, что этой особенности не было у Prestosuchus, которого он исключил из группы. Более поздние исследования обнаружили поддержку размещения Prestosuchus глубоко в Сучии, поэтому синапоморфия Серено, вероятно, недействительна.
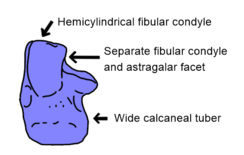 Левая пяточная кость (пяточная кость) Batrachotomus, демонстрирующая некоторые из пяточных особенностей Nesbitt (2011), которые считаются синапоморфными для Сухии
Левая пяточная кость (пяточная кость) Batrachotomus, демонстрирующая некоторые из пяточных особенностей Nesbitt (2011), которые считаются синапоморфными для Сухии Основной анализ ранних архозавров Стерлингом Несбиттом в 2011 году обнаружил несколько более стабильных сиапоморфий Сухии. Скуловые (скулы, ниже глаз) имеют низкую, закругленный продольный гребень в suchians, контрастирующая с другими археозаврами, которые, как правило, не имеют гребня, или острый гребень вместо. Как и в случае со многими ранними архозаврами, структура пяточной кости (наружная пяточная кость) важна для диагностики сучии. Эта кость соединяется как с малоберцовой костью (наружная кость голени), так и с астрагалом (внутренней костью лодыжки) с помощью двух различных поверхностей: выпуклого мыщелка малоберцовой кости и вогнутой астрагаларной фасетки. У других архозавров эти поверхности являются непрерывными, а не отдельными. Более того, мыщелок малоберцовой кости имеет полуцилиндрическую форму (форму полуцилиндра) в отличие от более сферической структуры орнитозухид, авеметатарсалий и родственников архозавров, таких как Euparkeria и proterochampsians. Полуцилиндрический мыщелок малоберцовой кости также присутствует у фитозавров, скорее всего, конвергентно. Наконец, задний край пяточной кости имеет проходящую назад костную трубку, известную как пяточный бугорок. У ранних архозавров пяточный бугорок был выше ширины, тогда как более поздние архозавры и родственники архозавров имели пяточный бугорок круглой формы в поперечном сечении. Такие люди пошли еще дальше, так как у ранних членов группы были пяточные клубни, которые были намного шире, чем высокие. Хотя двуногие попозавроиды и большинство лорикатанов ( Luperosuchus и корона впереди ) имели круглые пяточные клубни, эти особенности, вероятно, были реверсией, поскольку самые ранние попозавроиды ( например, Qianosuchus ) и лорикатаны ( Batrachotomus ) сохраняли широкие пяточные клубни.
У некоторых сучков есть неглубокая продольная бороздка на заднем крае лучевой кости (внутренняя кость предплечья), хотя распространение этой характеристики плохо изучено. Точно так же у некоторых ранних сицианцев был треугольный медиальный мыщелок бедренной кости (бедренной кости), но эта характеристика также известна у некоторых родственников архозавров, поэтому неясно, было ли наличие этого признака у такихианцев синапоморфией или плезиоморфией («примитивность»). черта, унаследованная от более раннего предка). Кроме того, большинство лорикатанов вернулись к состоянию, присущему другим архозаврам: округлому медиальному мыщелку. Многие из таких синапоморфий Несбитта (2011) отсутствуют у Nundasuchus, необычного круротарсана, который иногда считают таковым, близким к Ticinosuchus и Paracrocodylomorpha.
 Череп грацилизуха. Обратите внимание на закругленный гребень на скуловой кости под глазом (одна из синапоморфий Несбитта [2011]), видимый на A, и постеродорсальный отросток верхней челюсти (одна из синапоморфий Ezcurra [2016]), видимый на E ( pdpm)
Череп грацилизуха. Обратите внимание на закругленный гребень на скуловой кости под глазом (одна из синапоморфий Несбитта [2011]), видимый на A, и постеродорсальный отросток верхней челюсти (одна из синапоморфий Ezcurra [2016]), видимый на E ( pdpm) Анализ архозавроморфов, проведенный Мартином Эскуррой в 2016 году, предоставил альтернативный диагноз для Сучии. Эскурра обнаружил, что Койламасухус был членом группы, но также утверждал, что Сучия исключил Нундасухуса. В результате его диагноз Сухии зависел в первую очередь от нескольких черт, видимых у Koilamasuchus, который был известен по значительно меньшему количеству ископаемых материалов, чем большинство других Сухий. Эти признаки включают наличие больших ямок с хорошо окаймленными краями по обе стороны от спинных (задних) позвонков, плечевой кости ( плечевой кости), которая имеет симметричную проксимальную часть, если смотреть спереди, и преацетабулярного отростка (передняя лопатка). из подвздошной кости (верхние кости бедра), который имеет умеренную длину, длиннее, чем она высока, но не больше, чем лобковое стебло. Он также отметил, что у некоторых сабихов кончик заднего отростка верхнечелюстной кости (задняя ветвь) рядом с задним нижним кончиком анторбитального отверстия на самом деле выше средней части задней ветви. Эта черта, официально известная как постеродорсальный отросток верхней челюсти, считается синапоморфией грацилисухид и также присутствует у этозавров и Qianosuchus. Несмотря на широкое распространение этой характеристики, ее отсутствие у некоторых таких особей делает неоднозначным, квалифицируется ли она как синапоморфия Сухии, или, наоборот, эволюционировала в несколько независимых ветвей внутри группы. Эскурра также согласился с одной из синапоморфий пяточной кости Несбитта (2011), а именно, что пяточный бугорок шире, чем высокий.
Первоначально сухия была формализована немецким палеонтологом Бернардом Кребсом в 1974 году. Его намерением было связать триасовых псевдозухий (которые в то время считались не оставившими потомков) с настоящими крокодилами, которые эволюционировали позже в мезозое. Suchia был назван как порядок объединяет владение crurotarsal лодыжки, с Pseudosuchia и Крокодилами в качестве независимого подотряда внутри него. Однако более новые исследования, начатые в 1980-х годах, значительно изменили традиционную схему классификации предков крокодилов. Вместо того, чтобы быть родственными группами псевдозухий и крокодилов, как полагал Кребс, крокодилы теперь считаются подмножеством псевдозухий. Это потребовало от Сучиа нового определения, поскольку первоначальная классификационная схема Креба была признана недействительной.
Серено (1991) присвоил название Сучия кладе, содержащей большинство псевдозухий, которые ранее были обнаружены Бентоном и Кларком (1988). Согласно этому новому определению, Сухия включала Gracilisuchus stipanicicorum, Aetosauria, « Rauisuchia », Poposauridae, Crocodylomorpha и всех потомков их общего предка. Несбитт (2011) немного изменил определение Серено, поскольку он определил Сусию как наименее инклюзивную кладу, содержащую Aetosaurus ferratus, Rauisuchus tiradentes, Prestosuchus chiniquensis и Crocodylus niloticus (живого нильского крокодила ). Сучия, таким образом, представляет собой таксон на основе узлов, включающий крокодиломорфы и еще несколько базальных триасовых псевдозухих групп. К таким группам относятся Gracilisuchidae, Revueltosaurus, Aetosauria, Ticinosuchus, Poposauroida, Loricata и, возможно, Erpetosuchidae, Koilamasuchus, Nundasuchus и / или Euscolosuchus.
Таксоны, используемые Несбиттом (2011) для определения Сучии
Аэтозавр феррат
Rauisuchus tiradentes
Prestosuchus chiniquensis

Crocodylus niloticus
Ниже представлена филогенетическая кладограмма Батлера и соавт. в 2011 году демонстрирует кладистику Archosauriformes, уделяя основное внимание псевдозухиям : названия Clade следуют за Nesbitt 2011.
| Круротарси |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||