
Войти
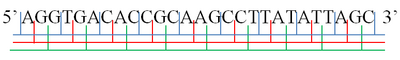 Иллюстрация возможных рамок чтения: AGG · TGA · CAC · CGC · AAG · CCT · TAT · ATT · AGC . A· GGT · GAC · ACC · GCA · AGC · CTT · ATA · TTA · GC. AG · GTG · ACA · CCG · CAA · GCC · TTA · TAT · TAG · C
Иллюстрация возможных рамок чтения: AGG · TGA · CAC · CGC · AAG · CCT · TAT · ATT · AGC . A· GGT · GAC · ACC · GCA · AGC · CTT · ATA · TTA · GC. AG · GTG · ACA · CCG · CAA · GCC · TTA · TAT · TAG · CВ молекулярной биологии рамка считывания представляет собой способ деления последовательности нуклеотидов в нуклеиновой кислоте (ДНК или РНК ) в набор последовательных, неперекрывающихся триплетов. Если эти триплеты приравниваются к аминокислотам или стоп-сигналам во время трансляции, они называются кодонами.
. Одна цепь молекулы нуклеиновой кислоты имеет фосфорильный конец, называемый 5'-концом, и гидроксил или 3'-конец. Они определяют направление 5 ′ → 3 ′. Есть три рамки считывания, которые могут быть прочитаны в этом направлении 5 '→ 3', каждая из которых начинается с другого нуклеотида в триплете. В двухцепочечной нуклеиновой кислоте дополнительные три рамки считывания могут считываться с другой, комплементарной цепи в направлении 5 '→ 3' вдоль этой цепи. Поскольку две цепи двухцепочечной молекулы нуклеиновой кислоты антипараллельны, направление 5 '→ 3' на второй цепи соответствует направлению 3 '→ 5' вдоль первой цепи.
В общем, при в наибольшей степени одна рамка считывания в данном разделе нуклеиновой кислоты является биологически релевантной (открытая рамка считывания ). Некоторые вирусные транскрипты можно переводить с использованием нескольких перекрывающихся рамок считывания. Существует один известный пример перекрывания рамок считывания в митохондриальной ДНК млекопитающих: кодирующие части генов для 2 субъединиц АТФазы перекрываются.
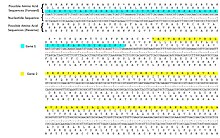 Пример трансляции с шестью кадрами
Пример трансляции с шестью кадрами ДНК кодирует последовательность белка серией трехнуклеотидных кодонов . Следовательно, любую данную последовательность ДНК можно прочитать шестью разными способами: три рамки считывания в одном направлении (начиная с разных нуклеотидов) и три в противоположном направлении. Во время транскрипции РНК-полимераза считывает цепь ДНК-матрицы в направлении 3 '→ 5', но мРНК образуется в направлении от 5 'к 3'. МРНК является одноцепочечной и поэтому содержит только три возможных рамки считывания, из которых только одна транслируется. Кодоны рамки считывания мРНК транслируются в направлении 5 '→ 3' в аминокислоты с помощью рибосомы с образованием полипептидной цепи.
Открытая рамка считывания (ORF) представляет собой рамку считывания, которая потенциально может быть транскрибирована в РНК и транслирована в белок. Для этого требуется непрерывная последовательность ДНК от стартового кодона через последующую область, которая обычно имеет длину, кратную 3 нуклеотидам, до стоп-кодона в той же рамке считывания..
Когда предполагаемая аминокислотная последовательность, полученная в результате трансляции ORF, оставалась неизвестной в митохондриальном и хлоропластном геномах, соответствующая открытая рамка считывания была названа неидентифицированной рамкой считывания (URF). Например, ген MT-ATP8 был впервые описан как URF A6L, когда был секвенирован полный митохондриальный геном человека.
 Два считывания рамки, используемые митохондриальными генами человека MT-ATP8 и MT-ATP6.
Два считывания рамки, используемые митохондриальными генами человека MT-ATP8 и MT-ATP6. Использование нескольких рамок считывания приводит к возможности перекрытия генов ; их может быть много в вирусных, прокариотных и митохондриальных геномах. Некоторые вирусы, например вирус гепатита B и BYDV, используют несколько перекрывающихся генов в разных рамках считывания.
В редких случаях рибосома может перемещаться из одного кадра в другой во время трансляции мРНК (сдвиг рамки трансляции ). Это приводит к тому, что первая часть мРНК транслируется в одной рамке считывания, а вторая часть транслируется в другой рамке считывания. Это отличается от мутации сдвига рамки считывания, поскольку нуклеотидная последовательность (ДНК или РНК) не изменяется - изменяется только рамка, в которой она считывается.