
Войти
| Вирус гепатита B | |
|---|---|
 | |
| Просвечивающая электронная микроскопия микрофотография показывает вирус гепатита B вирионы | |
| Классификация вируса | |
| (без рейтинга): | Вирус |
| Область: | Рибовирия |
| Королевство: | Парарнавиры |
| Тип: | Artverviricota |
| Класс: | Revtraviricetes |
| Порядок: | Blubervirales |
| Семейство: | Hepadnaviridae |
| Род: | Orthohepadnavirus |
| Виды: | вирус гепатита B |
Вирус гепатита B (HBV ), представляет собой частично двухцепочечный ДНК-вирус , вид рода Orthohepadnavirus и член Семейство вирусов Hepadnaviridae. Этот вирус вызывает заболевание гепатит B.
. Несмотря на то, что существует вакцина для предотвращения гепатита B, HBV остается глобальной проблемой здравоохранения. Гепатит B может быть острым, а затем перейти в хронический, приводя к другим заболеваниям и состояниям здоровья. Помимо гепатита, инфекция HBV может привести к циррозу и гепатоцеллюлярной карциноме.
Также было высказано предположение, что это может увеличить риск рака поджелудочной железы.
Вирусная инфекция вирусом гепатита B (HBV) вызывает множество изменений гепатоцитов из-за прямого действия белка, кодируемого вирусом, HBx, и непрямого изменения из-за значительного увеличения внутриклеточных активных форм кислорода (ROS) после заражения. HBx, по-видимому, нарушает регуляцию ряда клеточных путей. HBx частично вызывает нарушение регуляции за счет связывания с геномной ДНК, изменения паттернов экспрессии miRNA, воздействия на гистоновые метилтрансферазы, связывания с белком SIRT1 для активации транскрипции и взаимодействуя с гистон метилазами и деметилазами для изменения паттернов клеточной экспрессии. HBx частично ответственен за примерно 10 000-кратное увеличение внутриклеточных ROS при хронической инфекции HBV. Повышение АФК может быть частично вызвано локализацией HBx в митохондриях, где HBx снижает потенциал митохондриальной мембраны. Кроме того, другой белок HBV , HBsAg, также увеличивает АФК за счет взаимодействия с эндоплазматическим ретикулумом.
Увеличение активных форм кислорода (АФК) после инфицирования HBV вызывает воспаление, что приводит к дальнейшему увеличению ROS. АФК вызывают более 20 типов повреждений ДНК. Окислительное повреждение ДНК является мутагенным. Кроме того, восстановление повреждения ДНК может вызвать эпигенетические изменения в месте повреждения во время восстановления ДНК. Эпигенетические изменения и мутации могут вызвать дефекты в клеточном механизме, которые затем способствуют заболеванию печени.. К тому времени, когда накапливающиеся эпигенетические и мутационные изменения в конечном итоге вызывают прогрессирование до рака, эпигенетические изменения, по-видимому, играют большую роль в этом канцерогенезе, чем мутации. Только один или два гена, TP53 и, возможно, ARID1A, мутируют более чем в 20% раковых заболеваний печени, в то время как каждый из 41 гена имеет гиперметилированные промоторы (подавление экспрессии гена) более чем в 20% случаев рака печени, причем семь из этих генов гиперметилированы более чем в 75% случаев рака печени. Помимо изменений в участках репарации ДНК, эпигенетические изменения также вызываются HBx, рекрутирующим ферменты ДНК-метилтрансфераза , DNMT1 и / или DNMT3A., к конкретным локусам генов для изменения их уровней метилирования и экспрессии генов. HBx также изменяет ацетилирование гистонов, что может влиять на экспрессию генов.
Несколько тысяч генов, кодирующих белок, по-видимому, имеют сайты связывания HBx. Помимо генов, кодирующих белок, примерно 15 микроРНК и 16 длинных некодирующих РНК также подвержены влиянию связывания HBx с их промоторами. Каждая измененная микроРНК может влиять на экспрессию нескольких сотен информационных РНК (см. микроРНК ).
Вирус гепатита B классифицируется как типовой вид из ортогепаднавируса, который включает восемь других видов. Род классифицируется как часть семейства Hepadnaviridae, которое включает еще один род, Avihepadnavirus. Это семейство вирусов не было отнесено к вирусному ряду. Вирусы, подобные гепатиту B, были обнаружены у всех обезьян (орангутаны, гиббоны, гориллы и шимпанзе ), в обезьянах Старого Света и в Новом Свете шерстистых обезьянах (вирус гепатита В шерстистых обезьян ), что свидетельствует о древнем происхождении этот вирус у приматов.
Вирус разделен на четыре основных серотипа (adr, adw, ayr, ayw) на основе антигенных эпитопов, присутствующих на его белках оболочки. Эти серотипы основаны на общем детерминанте (а) и двух взаимоисключающих парах детерминант (d / y и w / r). Вирусные штаммы также были разделены на десять генотипов (A – J) и сорок субгенотипов в соответствии с общими вариациями нуклеотидной последовательности генома. Генотипы имеют четкое географическое распределение и используются для отслеживания эволюции и передачи вируса. Различия между генотипами влияют на тяжесть заболевания, течение и вероятность осложнений, а также реакцию на лечение и, возможно, вакцинацию. Серотипы и генотипы не обязательно совпадают.
Генотип D состоит из 10 субгенотипов.
Ряд еще не классифицированных видов, подобных гепатиту B, выделен от летучих мышей.
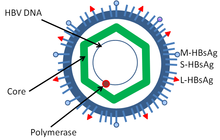 Структура вируса гепатита B
Структура вируса гепатита B Вирус гепатита B является членом семейства гепаднавирусов. Вирусная частица, называемая частицей Дейна (вирион ), состоит из внешней липидной оболочки и икосаэдрической ядра нуклеокапсида, состоящей из белок. Нуклеокапсид включает вирусную ДНК и ДНК-полимеразу, которая имеет активность обратной транскриптазы, аналогичную ретровирусам. Внешняя оболочка содержит встроенные белки, которые участвуют в связывании вируса и проникновении в чувствительные клетки. Вирус является одним из самых маленьких вирусов животных в оболочке с диаметром вириона 42 нм, но существуют плеоморфные формы, включая нитчатые и сферические тела без ядра. Эти частицы не являются инфекционными и состоят из липидов и белков, которые образуют часть поверхности вириона, называемого поверхностным антигеном (HBsAg ), и вырабатываются в избытке в течение жизненного цикла
Он состоит из:
Вирус гепатита D требует, чтобы частицы оболочки HBV стали вирулентными.
Трудно установить раннюю эволюцию гепатита B, как и всех других вирусов.
Дивергенция ортогепаднавируса и авихепаднавируса произошла ~ 125 000 лет назад (95% интервал 78 297–313 500). Вирусы Avihepadnavirus и Orthohepadna начали диверсифицироваться около 25 000 лет назад. Ветвление в это время привело к появлению генотипов Orthohepadna A – H. У человеческих штаммов самый недавний общий предок датируется от 7000 (интервал 95%: 5 287–9 270) до 10 000 (интервал 95%: 6 305–16 681) лет назад.
В авихепаднавирусе отсутствует Х-белок, но в геноме гепаднавируса уток присутствует рудиментарная рамка считывания Х. Белок X, возможно, произошел из ДНК-гликозилазы.
. Скорость несинонимичных мутаций в этом вирусе, по оценкам, составляет примерно 2 × 10 замен аминокислот на сайт в год. Среднее количество замен нуклеотидов на сайт в год составляет ~ 7,9 × 10.
Вторая оценка происхождения этого вируса предполагает, что он был самым недавним общим предком человеческих штаммов, появившихся около 1500 лет назад. Самый недавний общий предок птичьих штаммов был установлен 6000 лет назад. Частота мутаций составила ~ 10 замен на сайт в год.
Другой анализ с большим набором данных предполагает, что гепатит B инфицировал людей 33 600 лет назад (на 95% выше задняя плотность 22 000-47 100 лет назад. Расчетная скорость замещения составляла 2,2 × 10 замен на сайт в год. рост популяции был отмечен в течение последних 5000 лет. Межвидовое заражение орангутангов и гиббонов произошло в течение последних 6100 лет.
Изучение последовательностей у зебровых вьюрков отодвинуло происхождение этого рода, по крайней мере, до 40 миллионов лет назад и, возможно, до 80 миллионов лет назад. Шимпанзе, горилла, орангутанг и гиббоны Кластер видов с человеческими изолятами. Среди неприматов были вирус гепатита сурка, вирус гепатита суслика и вирус гепатита арктической белки. Также был описан ряд видов, заражающих летучих мышей. Было высказано предположение, что летучие мыши Нового Света могут быть происхождение вида приматов.
Исследование изолятов f Из циркумполярной арктической популяции людей было высказано предположение, что предок субгенотипа B5 (эндемический тип, обнаруженный в этой популяции), что предковый вирус возник в Азии около 2000 лет назад (95% HPD 900 г. до н.э. - 830 г. н.э.). Слияние произошло около 1000 г. н.э. Этот субгенотип сначала распространился из Азии в Гренландию, а затем за последние 400 лет распространился на запад.
Самые старые свидетельства инфекции гепатита В относятся к бронзовому веку. Доказательства были получены из человеческих останков возрастом 4500 лет. Согласно исследованию 2018 года, вирусные геномы, полученные с помощью секвенирования дробовика, стали самыми старыми из когда-либо извлеченных из образцов позвоночных. Также было обнаружено, что некоторые древние штаммы вируса гепатита все еще инфицируют людей, а другие вымерли. Это опровергло мнение о том, что гепатит В возник в Новом Свете и распространился в Европу примерно в 16 веке.
 Организация генома HBV. Гены перекрываются.
Организация генома HBV. Гены перекрываются. Геном HBV состоит из кольцевой ДНК, но это необычно, потому что ДНК не является полностью двухцепочечной. Один конец полноразмерной цепи связан с вирусной ДНК-полимеразой. Геном имеет длину 3020–3320 нуклеотидов (для полной цепи) и 1700–2800 нуклеотидов (для короткой цепи).
Отрицательный -смысл, (некодирующая) цепь комплементарна вирусной мРНК. Вирусная ДНК обнаруживается в ядре вскоре после заражения клетки. Частично двухцепочечная ДНК становится полностью двухцепочечной за счет завершения (+) смысловой цепи клеточными ДНК-полимеразами (вирусная ДНК-полимераза используется на более поздней стадии) и удалением вирусного полимеразного белка ( P) из (-) смысловой цепи и короткой последовательности РНК из (+) смысловой цепи. Некодирующие основания удаляются с концов (-) смысловой цепи, и концы соединяются заново.
Вирусные гены транскрибируются клеточной РНК-полимеразой II в ядре клетки из матрицы ковалентно замкнутой кольцевой ДНК (кзкДНК). В геноме HBV были идентифицированы два энхансера, обозначенные как энхансер I (EnhI) и энхансер II (EnhII). Оба энхансера проявляют большую активность в клетках печеночного происхождения, и вместе они управляют и регулируют экспрессию полных вирусных транскриптов. Существует четыре известных гена, кодируемых геномом, которые называются C, P, S и X. Основной белок кодируется геном C (HBcAg), и его стартовому кодону предшествует начальный кодон AUG в рамке считывания., из которого производится предварительный коровый белок. HBeAg продуцируется протеолитическим процессингом пре-корового белка. ДНК-полимераза кодируется геном P. Ген S - это ген, кодирующий поверхностный антиген (HBsAg). Ген HBsAg представляет собой одну длинную открытую рамку считывания, но содержит три кодона в «стартовой» рамке (ATG), которые делят ген на три участка: пре-S1, пре-S2 и S. Из-за множественных стартовых кодонов полипептиды трех производятся разные размеры, называемые большим, средним и малым (pre-S1 + pre-S2 + S, pre-S2 + S или S). Функция белка, кодируемого геном X, до конца не изучена, но некоторые данные свидетельствуют о том, что он может функционировать как транскрипционный трансактиватор.
Несколько элементов некодирующей РНК были идентифицированы в геноме HBV. К ним относятся: HBV PREalpha, HBV PREbeta и сигнал инкапсидации РНК HBV эпсилон.
Генотипы отличаются не менее чем на 8% последовательности и имеют четкое географическое распространение, и это было связано с антропологической историей. Внутри генотипов были описаны подтипы: они различаются на 4–8% генома.
Существует восемь известных генотипов, помеченных от A до H.
Был описан возможный новый генотип «I», но принятие этой нотации не является универсальным.
С тех пор были распознаны еще два генотипа. В текущем (2014 г.) листинге от A до J. Также распознаются несколько подтипов.
Есть как минимум 24 подтипа.
Различные генотипы могут реагировать на лечение по-разному.
Тип F, который отличается от других геномов на 14%, является наиболее дивергентным из известных типов. Тип A распространен в Европе, Африке и Юго-Восточной Азии, включая Филиппины. Типы B и C преобладают в Азии ; тип D распространен в районе Средиземноморья, Ближнего Востока и Индии ; тип E локализован в Африке к югу от Сахары; тип F (или H) ограничен Центральной и Южной Америкой. Тип G был обнаружен в Франции и Германии. Генотипы A, D и F преобладают в Бразилии, и все генотипы встречаются в США с частотами, зависящими от этнической принадлежности.
Штаммы E и F, по-видимому, произошли от аборигенных популяций Африки и Нового Света соответственно.
Тип A имеет два подтипа: Aa (A1) в Африке / Азии и на Филиппинах и Ae (A2) в Европе / США.
Тип B имеет два различных географических распределения: Bj / B1 ('j' - Япония) и Ba / B2 ('a' - Азия). Тип Ba был далее подразделен на четыре клады (B2 – B4).
Тип C имеет два географических подтипа: Cs (C1) в Юго-Восточной Азии и Ce (C2) в Восточной Азии. Подтипы C были разделены на пять клад (C1 – C5). Шестая клада (C6) была описана на Филиппинах, но на сегодняшний день только в одном изоляте. Тип C1 связан с Вьетнамом, Мьянмой и Таиландом ; введите C2 с Япония, Корея и Китай ; введите C3 с Новой Каледонией и Полинезией ; C4 с Австралией ; и C5 с Филиппинами. Другой подтип был описан в Папуа, Индонезия.
Тип D был разделен на 7 подтипов (D1 – D7).
Тип F разделен на 4 подтипа (F1 – F4). F1 был дополнительно разделен на 1a и 1b. В Венесуэле подтипы F1, F2 и F3 встречаются у индейцев Востока и Запада. Среди южноамериканских индейцев был обнаружен только F3. Подтипы Ia, III и IV имеют ограниченное географическое распространение (Центральная Америка, Север и Юг Южной Америки соответственно), в то время как клады Ib и II встречаются во всех Америках, за исключением Северной Южной Америки и Северной Америки соответственно.
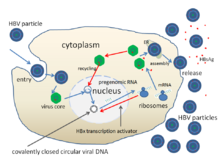 Репликация вируса гепатита B
Репликация вируса гепатита B Жизненный цикл вируса гепатита B сложен. Гепатит B - один из немногих известных неретровирусных вирусов, которые используют обратную транскрипцию как часть процесса репликации.
HBV обладают способностью трансактивировать FAM46A.