
Войти
 A жужжалица, у которой булавовидные жужжальцы видны непосредственно позади места прикрепления крыльев к телу.
A жужжалица, у которой булавовидные жужжальцы видны непосредственно позади места прикрепления крыльев к телу.  Жужжальца-краболова
Жужжальца-краболова  Воспроизвести медиа Жужжальца движущегося
Воспроизвести медиа Жужжальца движущегося Жужжальца (; единственное число недоуздок или жужжальца ) (от Древнегреческого : λτῆρες, гиря, удерживаемая в руках, чтобы дать толчок к прыжку) - пара маленьких булавовидных органов на теле двух Приказов полета насекомые, дающие информацию о поворотах тела во время полета. Примеры насекомых с жужжальцами: комнатные мухи, комары, мошки и журавлиные мухи.
Жужжальцы быстро колеблются вместе с крыльями и как гироскопы с вибрирующей структурой : любое вращающееся колебание вызывает вибрирующие жужжальцах за счет эффекта Кориолиса. Насекомое обнаруживает эту силу с помощью органов чувств, называемых колоколообразными сенсиллами и хордотон примени, расположенными у основания жужжальцев, и использует эту информацию для интерпретации и корректировки своего положения в пространстве. Жужжальца обеспечивают быструю обратную связь с мускулами, управляющими крыльями, а также с мышцами, отвечающими за стабилизацию головы.
У насекомых большого отряда Diptera (мухи) есть жужжальца, которые произошли от насекомых. пара предковых задних крыльев, в то время как самцы гораздо меньшего отряда Strepsiptera (стилопы) имеют жужжальца, которые произошли от пары предковых передних крыльев.
У насекомых две пары крыльев. У мух есть только один набор крыльев, создающих подъемную силу, и один набор жужжалок. Название отряда мух, «Diptera», буквально означает «два крыла», но есть другой отряд насекомых, которые развили полет только с двумя крыльями: strepsipterans или стилопсы; они единственные другие организмы, у которых есть два крыла и два жужжальца. Стреперы приспособили свои передние крылья в жужжальца, тогда как двукрылые превратили свои задние крылья в жужжальцев. Эта уникальная структура, которая создает вращения / возмущения во время полета, никогда не была описана в природе где-либо еще.
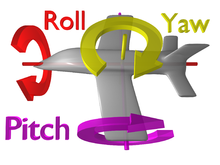 Направления вращения
Направления вращения Жужжальца способны проявлять небольшие отклонения в положении тела, используя гироскопические свойства движущейся массы.. Это означает, что жужжальцы бьются вверх и вниз в такт с взмахом крыльев по линейному пути, но когда тело мухи начинает вращаться, траектория бьющихся жужжальцев также меняется. Теперь вместо жужжальцев, они начинают следовать по кривой. Чем сильнее возмущение, которое они испытывают, тем дальше жужжальцы отклоняются от своего первоначального линейного пути. В эти периоды жужжальца движутся не только в двух направлениях (вверх и вниз), а в четырех (вверх, вниз, влево и вправо). Сила, действующая на жужжальца в ответ на это движение влево-вправо, известна как сила Кориолиса и может создаваться, когда любой движущийся объект вращается в трех направлениях: вращение, рыскание, тангаж. или свернуть (см. рисунок). Когда это происходит, крошечные колоколообразные структуры у основания жужжальца испытывают напряжение, поскольку стебель жужжальца изгибается в их направлении. Затем нервная система может преобразовать изгиб этих волосков в электрические сигналы, которые муха интерпретирует как информацию о вращении тела. Муха использует эту информацию, чтобы корректировать положение во время полета. Дальнейшие подробности, объясняющие динамику и физиологию жужжальцев, рост ниже.
Зужжальца обычно связаны только со стабилизацией, но их способность обнаруживать повороты тела может вызывать компенсаторные механизмы не только со стороны управляющих мышц крыльев, но также со стороны шейных мышц, которые соответствуют положению головы и взгляд. Жужжальцы также могут быть полезны для другого поведения. У некоторых видов мух было замечено, что они раскачивают жужжальца во время ходьбы в дополнение к колебаниям во время полета. Таким образом, у этих людей жужжальцы могут обнаруживать сенсорную информацию и во время ходьбы. Когда жужжальца удалены, эти насекомые хуже справляются с некоторыми трудностями при ходьбе. Однако, как обрабатывается и используется жужжалочная информация во время ходьбы, остается, за некоторыми исключениями, неясным. Конкретные примеры того, что было обнаружено, возможности ниже.
Жужжальца были впервые задокументированы Уильямом Дерхамом в 1714 году. Он обнаружил, что мухи не могут оставаться в воздухе, когда их жужжальцы были удалены хирургическим путем, но в остальном ведут себя нормально. Первоначально этот результат был приписан способности жужжальца поддерживать равновесие. В 1917 г. фон Бадденброк утверждал, что-то еще было причиной потери мухами способности летать. Он утверждал, что вместо этого жужжальца следует рассматривать как «стимулирующие органы». Другими словами, деятельность жужжальцев приводила в действие мышечную систему крыла, так что они действовали как переключатель включения / выключения для полета. В. Будденброк попытался показать, что активация жужжальцев будет стимулировать центральную нервную систему в таком состоянии активности, позволяет крыльям выполнять полетное поведение. С тех пор был сделан вывод, что это на самом деле неправда, и что это настоящее утверждение, что жужжальцы как органы равновесия, является правильным. В. Будденброку удалось показать, что сразу после удаления жужжальца мухи не могут быть нормальные движения крыльев. Позже это было объяснено тем фактом, предоставление мухам времени на восстановление за несколько минут после операции к полному восстановлению нормального контроля над мышцами полета. Кроме того, в интересном побочном эксперименте, проведенном Принглом (1938), когда нить прикрепляли к брюшку беспорванных мух, снова достигалась стабильная полётка. Нить в этих экспериментах предположительно помогла мухе не вращаться (как тяжелая корзина под воздушным шаром предотвращает опрокидывание шара), что подтверждает гипотезу о том, что жужжальцы ответственны за восприятие тела.
Первоначальная теория балансира, постулированная Принглом (1948), учитывала только силы, учитывающие в двух направлениях. Прингл утвержден, что рыскание было единственным направлением вращения, которое мухи использовали для обнаружения своих жужжальцев. Используя высокоскоростную видеоанализ, «Фауст» (1952) показал, что это не так, и что жужжальцы способны обнаруживать все направления вращения. В ответ на это новое открытие Прингл пересмотрел свое предыдущее предположение и пришел к выводу, что мухи показали все три направления вращения, просто сравнивая сигналы с левой и правой сторон тела. Конечно, это не настоящий механизм, с помощью которого мухи обнаруживают вращение. Различные поля органов, расположенных в разных областях у основания жужжальца, определяют разные направления вращения, которые также объясняют, почему мухи с одним жужжальцем все еще летать без проблем.
Принято считать, что жужжальцы произошли от нелетных крыльев насекомых - задних крыльев двукрылых и передних крыльев Strepsiptera соответственно. Их движение, структура, функции и развитие подтверждают эту гипотезу. Характеристики органов чувств, известные как колоколообразные сенсиллы, обнаруженные в основании жужжальца, показывают много сходства с таковыми у других насекомых у основания задних крыльев. Сенсиллы устроены так, как у задних крыльев, что если бы жужжальца были заменены крыльями, создаваемые сил было достаточно для активации тех же органов чувств. Генетические исследования также выявили много общего между жужжальцами и задними крыльями. Фактически, развитие жужжальцы восходит к одному гену (Ubx), который при деактивации приводит к образованию заднего крыла. Это изменение отвечает только один ген, легко представить себе небольшую мутацию, приводящую к созданию первого жужжалок.
Хотя никакой другой структуры с полностью в природе наблюдались те же функции и морфология, что и у жужжальцев, они развивались как минимум дважды в классе Насекомые, раз в порядке Diptera и еще раз в Strepsiptera. В классе насекомых также существует другая структура, основная функция которой не такая функция, как у жужжальцев. Это происходит в отряде Lepidoptera и относится к усикам мотыльков и бабочек.
 Стрепсиптерановые жужжальцы: каплевидные структуры, расположенные между передними и средними ногами (обозначенными стрелками) [2]
Стрепсиптерановые жужжальцы: каплевидные структуры, расположенные между передними и средними ногами (обозначенными стрелками) [2] Strepsipterans - уникальная группа насекомых с большим половым диморфизмом. Самки проводят свою жизнь в личинковом состоянии, паразитируя на более крупных насекомых. Единственный раз, когда они выходят из своего насекомого-хозяина, - это вытягивать сросшиеся головы и грудную клетку, чтобы самцы это заметили. Самцы тоже паразиты, но в итоге они покидают своего ина в поисках своих собратьев-самок. Из-за этого они все еще сохраняют способность летать. Стрепсиптеросые самцы однозначно обладают двумя задними крыльями, в то время как их передние крылья принимают форму булавы жужжальцев. Хотя стрептокрылых очень сложных конструкций и, кроме того, они довольно недолговечны, Pix et al. (1993) подтвердили, что специализированные передние крылья самцов Strepsiptera выполняют ту же функцию, что и двукрылые жужжальцы. Вращательные движения тела в сочетании с колеблющимися жужжальцами силы Кориолиса, которые могут быть обнаружены полями механодатчиков (колоколообразных сенсоров), применены в основании жужжальцев. Используя исследования функциональной морфологии и поведения, Pix et al. показывает, что эти датчики передают информацию о положении тела в голову и, чтобы произвести компенсирующие движения. Для простоты, оставшаяся часть этой статьи будет относиться только к двукрылым жужжальцам.
Некоторые чешуекрылые (мотыльки и бабочки) во время демонстрают колебания своей антенны с небольшой амплитудой под постоянными углами. Первоначально предполагалось, что движения антенн у чешуекрылых антенн в восприятии ветра или силы тяжести. Исследование, проведенное на ястребиной моли, Manduca sexta, подтвердило, что эти крошечные колебания антенн на самом деле способствуют возникновению ощущения вращения тела.
Sane et al. (2007) определили, что антенны были ответственны за стабилизацию стабилизации ястребиных бабочек, удалив длинную часть антенны (жгутик), а затем прикрепив ее, чтобы определить ее влияние на летные характеристики. Когда жгутики были удалены, бабочки больше не могли поддерживать стабильный полет. После повторного прикрепления жгутиков летные характеристики восстановились. Источник этой разницы был определен как механосенсорный. Есть два набора механосенсорных органов, установка у основания антенны чешуекрылых, щетинки Бема и орган Джонстона. Эти поля рецепторы реагируют на разные направления движения усиков. Усики также способны ощущать запах, влажность и температуру. Sane et al. (2007) показали, что именно механосенсоры были ответственны за стабильность в отличие от других органов, потому что, когда жгутики были удалены, а снова прикреплены, все усиковые нервы были разорваны, кроме тех, что у основания (щетина Бема и орган Джонстона).
В сегментированных организмах есть гены, называемые Hox-генами, которые определяют развитие серийных гомологов, или повторяющихся структур внутри организма (например, сочлененные отростки членистоногих или позвонков у млекопитающих). У насекомых грудная клетка разделена на разные сегменты. Одна из вещей, за которую отвечает ген Hox Ультрабиторакс (Ubx), - это определение идентичности грудного сегмента их тела. Правильное развитие задних крыльев у ряда видов насекомых зависит от Ubx, включая бабочек, жуков и мух. У плодовых мух (Ubx) отвечает за формирование жужжальцев во время метаморфоза. Если экспериментально деактивировать этот ген, жужжальца разовьется в полностью сформированное крыло. Это единственное изменение гомеотического гена приводит к радикально другому фенотипу, но также дает нам представление о том, как предки задних крыльев мух использовать развиться в жужжальцев.
Хотя очевидно, что Ubx является первичным геном, ответственным за формирование заднего крыла, Ubx также регулирует другие гены после экспрессии. Weatherbee (1998) предположил, что различия в паттернах или уровнях экспрессии Ubx не могут быть ответственны за наблюдаемые физиологические изменения. Вместо этого он предположил, что регулируемые наборы Ubx генов-мишеней были получены наблюдаемых изменений. Было идентифицировано несколько Ubx-регулируемых генов-мишеней, включая две прямые мишени, spalt и knot, которые экспрессируются в крыле и репрессируются в жужжальцах. Были идентифицированы и гены, которые экспрессируются в крыльях и репрессируются в других жужжальцах, но они в качестве прямых мишеней регуляции Ubx, пока неизвестно.
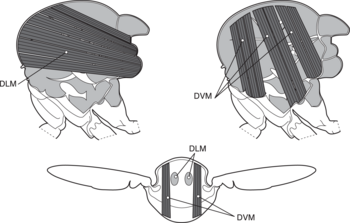 Грудная клетка мухи показывает вид сбоку на продольную продольную часть позвоночника. (DLM; вверху слева) и дорсо-вентральная (DVM; вверху справа) мышцы силового полета. На нижнем изображении показано поперечное сечение мухи [3].
Грудная клетка мухи показывает вид сбоку на продольную продольную часть позвоночника. (DLM; вверху слева) и дорсо-вентральная (DVM; вверху справа) мышцы силового полета. На нижнем изображении показано поперечное сечение мухи [3].Двуклонные насекомые вместе с большинством других отрядов используют так называемые мышцы непрямого полета для выполнения полета. Непрямые мышцы полета насекомых состоят из двух наборов перпендикулярных мышц (см. Левый рисунок), которые прикреплены к грудной клетке (а не непосредственно к основанию крыла, как в случае мышц прямого полета). Когда первый набор мышц сокращается, они деформируют насекомого и вертикально сжимают его грудную клетку, что поднимает крылья. Когда первая группа мышц расслабляется, а вторая сокращается, грудная клетка сжимается в противоположном направлении, расширяет тело вертикально и перемещает крылья вниз. На рисунке ниже показано это движение только с первым набором мышц.
 Движение крыла насекомого:. aкрылья. bпервичные и вторичные лётные суставы. cдорсовентральные лётные суставы. dпродольные мускулы
Движение крыла насекомого:. aкрылья. bпервичные и вторичные лётные суставы. cдорсовентральные лётные суставы. dпродольные мускулы Движение крыльев и жужжальцев механически связано. Sane et al. (2015) применимо, что у только что убитых мух без каких-либо нервных импульсов движение крыльев все еще было связано с движением жужжальцев. Когда для ручного перемещения крыла вверх и вниз использовался зажим, не только противоположное крыло двигалось синхронно, но и жужжальцы также ударялись в противофазе обоими крыльями. Однако источник этой связи был не между мускулами, управляющими жужжальцами, и мускулами, управляющими крыльями. Вместо этого были обнаружены два небольших гребня кутикулы, известные как субэпимерные гребни. Эти гребни соединяют правое крыло с правым недоуздком, а левое крыло с левым недоуздком.
Каждая сторона тела должна быть синхронизирована, и две стороны также должны быть соединены. То есть левое и правое крыло и, следовательно, левый и правый жужжальца всегда бьют с одинаковой величиной. Однако амплитуда взмаха крыльев не всегда должна быть одинаковой с левой и с правой стороны. Это то, что позволяет мухам поворачиваться, и достигается с помощью коробки передач, что вы найдете в автомобиле. Этот редуктор может увеличить максимальную амплитуду движения крыла и определить скорость его движения. Крылья мух даже имеют в основании сцепления. Муфта перемещается между канавками в коробке передач, чтобы задействовать и расцепить мышцы крыльев, а также изменить амплитуду биений крыльев. Когда амплитуда левого крыла меньше правого, муха сделает левый поворот. Несмотря на то, что движение жужжальца контролируется отдельными мышцами, а не крыльями, поскольку крылья механически связаны с жужжальцом, изменения в частотах взмахов крыльев распространяются также на частоту ударов жужжальца, но амплитуда ударов жужжальца не изменяются.
Хотя жужжальцы связаны с крыльями и друг с другом во время полета, некоторые мухи раскачивают свои жужжальцы при ходьбе (без раскачивания крыльев). Поскольку мускулы жужжальца крошечные по сравнению с мускулами полета, их активность полностью затмевает активность мускулов жужжальца во время полета. Неизвестно, чем активность мышц жужжальцы во время полета отличается от ходьбы. Левое и правое жужжальца демонстрируют гораздо более изменчивые фазовые отношения во время ходьбы по сравнению с полетом, что может указывать на разделение левой и правой мускулов жужжальца.
Хотя жужжальцы всегда синхронизированы с движения крыльев, фазы, в которых они колеблются, различаются у разных видов. Brachyceran мухи (с короткими усиками) колеблют свои жужжальцы почти точно напротив своих крыльев (180 градусов). Более древние подотряды, такие как Nematecerans (мухи с длинными усиками), к которым, например, относятся журавлиные мухи и комары, демонстрируют разнообразие фазировок крыла-недоуздка. Эти наблюдаемые различия в координации крыла и жужжальца указывают на то, что различия в продукции сенсорных нейронов также существуют между видами. Это означает, что механизмы декодирования, используемые центральной нервной системой для интерпретации таких движений и обеспечения адекватной двигательной активности, вероятно, также варьируются в зависимости от филогении.
 Электронная микрофотография жужжальца мухи и окружающих структуры:. 1calyptra (squama) 2 верхняя чашечка (антисквама) 3 haltere 4 mesopleuron 5 hypopleuron 6 тазик 7 крыло 8 брюшной сегмент 9 мезонотум c головная часть жужжальцы p ножка жужжальцы s чешуйка жужжальца
Электронная микрофотография жужжальца мухи и окружающих структуры:. 1calyptra (squama) 2 верхняя чашечка (антисквама) 3 haltere 4 mesopleuron 5 hypopleuron 6 тазик 7 крыло 8 брюшной сегмент 9 мезонотум c головная часть жужжальцы p ножка жужжальцы s чешуйка жужжальца Общая структура жужжальцев хорошо известна, но между видами существует много различий. Более древние семейства, такие как tipulidae (журавлиные мухи), обладают жужжальцами с довольно длинными стеблями. Это приводит к тому, что луковицы жужжальца находятся намного дальше от тела и хорошо видны невооруженным глазом. Более производные семейства, такие как Calliphoridae (летучие мухи), развили специализированные структуры, называемые «calyptrae» или «чешуйки», которые представляют собой крошечные крылья, покрывающие жужжальца. Прингл (1948) предположил, что они предотвращают влияние турбулентности ветра на движения жужжальца, позволяя более точно определять положение тела, но это никогда не было проверено. Стебель жужжальца тоже не всегда прямой. Вместо этого форма стебля в более производных семьях, как правило, отражает форму тела человека. Это минимизирует количество воздушного пространства между концевыми выступами и сторонами живота и грудной клетки. В этих семьях жужжальцы бьют так близко к телу, что расстояние между жужжальцами и грудной клеткой составляет долю диаметра луковицы жужжальца. Ярким примером этого признака является семейство Syrphidae (журчалки), где луковица жужжальца расположена почти перпендикулярно стеблю.
Мухи обычно держат жужжальца под углом 90 градусов. смещение. Чтобы визуализировать это, если вы представите человека, держащего руки встороны, это будет смещение на 180 градусов. Если бы этот человек переместил руки назад так, чтобы угол между кончиками пальцев и позвоночником составлял 90 градусов, это было бы смещением на 90 градусов. Жужжальца у мух работают точно так же. Они расположены позади своего тела, образуя угол в 90 градусов между луковицами жужжальцев и центром их грудной клетки. Необходимо, чтобы жужжальцы располагались таким образом, чтобы обнаруживать три оси движения. Эти оси представляют собой наклон и кренскания, как показано на рисунке выше (Направления вращения). механорецепторы в основании жужжальцев могут измерять силу только в двух направлениях (горизонтальном и вертикальном), поэтому один жужжальца может измерять вращение только по двум из трех осей. Временным жужжальцы установлены разными углами (смещение 90 градусов), они также бьют по горизонтальным и вертикальным осям. Это дает возможность получать информацию из двух непараллельных плоскостей и позволяет ощущать вращение во всех трех направлениях. Однако мухи наиболее чувствительны к смоле.
 Длина и форма жужжальца различного вида от вида к виду.
Длина и форма жужжальца различного вида от вида к виду. Когда жужжальца экспериментально заставляют взмахнуть, выстреливают залпами с потенциалом действия внутри нерва жужжальца работают синхронно с установкой ударов жужжальца. Когда мухи вращают, эти залпы разрушаются, вероятно, в ответ на активацию различных групп сенсилл, чтобы сообщить муху о ее недавнем положении тела. Было также показано, что афференты Haltere оканчиваются в мезоторакальным нейропиле, где расположены нейроны летательных мышц. Афферентная активность жужжальца, отвечающая на вращение и поведение крыльев, сходится в этой области обработки.
Сенсорные входы от пяти сенсорных полей, расположенных в основании жужжальца, все сходятся на одном нерве, нерве жужжальца. Как эти сенсорные поля организованы на уровне центральной нервной системы, в настоящее время неизвестно. Установлено, что все эти пять сенсорных полей проецируются на грудные клетку «регионально-специфическим», и афференты, исходящие от переднего крыла, также сходятся в одних и тех же регионах. Не все специфические мишени для афферентов жужжальца выявлены, но были идентифицированы несколько связей между моторными нейронами, которые, как известно, участвуют в управлении движением крыльев, определенными сенсорными полями жужжанки, в частности, одним синапс между нервными жужжальца и двигательный нейрон управления крыльями, известный как mnb1.
Мухи используют мышцы непрямого полета для выполнения движения крыльев, а биение жужжальца осуществляется той же группой мышц (см. Раздел динамики). В дополнение к мышцам непрямого полета, которые соответствуют взмахи, есть также рулевые мышцы, которые контролируют поворот / угол поворота крыльев. Указание на случай жужжальца происходит от задних крыльев. Чан и др. (1998) идентифицировали 10 мышц прямого управления в жужжальце, подобных тем, что обнаружены в переднем крыле. В 1998 году Чан и Дикинсон предположили, что запланированные движения на холтере (воздействия на них внешних сил) были тем, что инициировало запланированные повороты. Чтобы объяснить это, представьте муху, которая хочет повернуть направо. К сожалению, как только это происходит, жужжальцы улавливают вращение тела и рефлекторно корректируют поворот, не давая мухе изменить направление. Чан и Дикинсон (1998) предположили, что для предотвращения этого муха сначала перемещает жужжальца, как если бы ее толкали в направлении, противоположном ее желанию. Муха не двинулась с места, но жужжальцы почувствовали волнение. Это позволит проявиться рефлексу, вызванному жужжалкой, и исправить воображаемое возмущение. Тогда муха сможет выполнить свой поворот в желаемом направлении. На самом деле мухи летают не так. Мурели и Фокс (2015) показали, что мухи все еще способны выполнить запланированные повороты, даже если их жужжальца полностью удалены.
 Схема шести основных полей колокольчиков на жужжальце. Четыре поля установлены дорсально - дорсальные сосочки Хика (dHP), дорсальная базальная пластинка (dBP), дорсальная пластинка скапала (dSP) и дорсальная фланкирующая сенсилла (FS). Вентрально расположены два поля: вентральные сосочки Хика (vHP) и вентральная пластинка скапала (vSP).
Схема шести основных полей колокольчиков на жужжальце. Четыре поля установлены дорсально - дорсальные сосочки Хика (dHP), дорсальная базальная пластинка (dBP), дорсальная пластинка скапала (dSP) и дорсальная фланкирующая сенсилла (FS). Вентрально расположены два поля: вентральные сосочки Хика (vHP) и вентральная пластинка скапала (vSP). Способ, обеспечивающее ощущение вращения, заключается в том, что существует пять различных сенсорных полей, расположенных у основания жужжальца. Эти поля, которые на самом деле обеспечивают большинство колоколообразных сенсилл, обнаруженных на экзоскел мясных мух (более 400 колоколоных сенсилл на жужжальце), активируются в ответ на напряжение, создаваемое движениеми в основе недоуздка в разных направленийх (из-за сил Кориолиса, действующие на концевые ручки). Колоколообразные сенсиллы - это шапковидные выступы, расположенные на поверхности экзоскелета (кутикулы) насекомых. Внутри колпачка прикреплен конус дендритного выступа (или сенсорного волокна нервов). Внешний дендритного выступа прикреплен к внутренней поверхности шляпки. Когда жужжальца отодвигают в сторону, кутикула насекомого искривляется и поверхность шляпки искажается. Внутренний дендрит распознает и преобразовывает его в электрический сигнал, который отправляется в центральную нервную систему для интерпретации.
Хордотональные органы обнаруживают и передают искажения в своем положении / форме так же, как и у колоколообразных сенсилл. Они немного отличаются в месте обнаружения. Хордотональные органы, в отличие от колоколообразных сенсилл, находятся под кутикулой и обычно реагируют на растяжение, а не на деформацию или изгиб. Их сенсорные нервные окончания прикрепляются между двумя внутренними точками, и когда эти точки растягиваются, разница в длине - это то, что обнаруживается и преобразуется в электрические сигналы. В основании жужжальца гораздо меньше хордотональных органов, чем колоколообразных сенсилл (порядка сотен), поэтому они менее важны для обнаружения и передачи информации о вращении движений жужжальца.
Глаза насекомых не могут двигаться независимо от головы. Чтобы мухи могли стабилизировать свое поле зрения, они должны отрегулировать положение всей своей головы. Сенсорные сигналы, устанавливаемые жужжальцами, определяют не только положение тела, но также и положение головы, которая может двигаться независимо от тела. Халтеры особенно полезны для обнаружения быстрых возмущений во время полета и реагируют только на угловые скорости (скорости вращения) выше определенного порога. Когда мухи фокусируются на объекте перед ними. Хенгстенберг (1988) обнаружил, что в направлении вращения крена способность мух положения головы в ответ на движение наблюдалась только при скорости выше 50 градусов в секунду, а их способность достигала максимума при 1500 градусах в секунду. Когда жужжальца были удалены у луковицы (чтобы сохранить неповрежденные органы у основания), способность мухи воспринимать креновые движения с высокими угловыми скоростями исчезла.
Жужжальца и зрение играют роль в стабилизации головы. Мухи также может выполнять компенсирующие движения головой для стабилизации зрения без использования жужжальцев. Когда поле зрения искусственно вращается вокруг мухи с меньшими угловыми скоростями, стабилизация головы все же происходит. Выходы стабилизации головы из-за одних только оптических входов реагируют медленнее, но также снимают дольше, чем выходы из-за холтерных входов. Из этого результата можно сделать вывод, что хотя жужжальца необходимы для обнаружения быстрых вращений, зрительная система сама умеет распознавать и корректировать более медленные движения тела. Таким образом, зрительная и механосенсорная (жужжальца) системы работают вместе для стабилизации поля зрения животного: во-первых, быстро реагируя на быстрые изменения (жужжальцы), поддерживая эту реакцию до тех пор, пока она не будет исправлена (зрение).
Мухи полагается как на визуальную информацию, исходящую от их сложных, так и на механическую информацию от их жужжальцев. Шерман и Дикинсон (2002) представила, что реакция жужжальцев и глазена на дополнительные скорости вращения. Реакция на вращение тела, проявляемая через зрительную систему, максимальна при малых скоростях и влиянии угловой скорости. Напротив, вращение тела, обнаруженное жужжальцами, наибольшие отклики при более высоких угловых скоростях и плохо по мере уменьшения скорости вращения. Интеграция этих двух отдельно настроенных датчиков позволяет мухам обнаруживать диапазон угловых скоростей во всех трех направлениях вращения.
Для изучения зрения мух были использованы два основных поля зрения, рисунок и фон. Фигуры - это объекты, на которых сфокусирована муха, а фон представляет все остальное. Когда жужжальца снимают с привязанных летающих мух, они все еще могут двигаться движущиеся фигуры, но им сложно стабилизировать движущийся фон. Если статическая фигура находится в поле зрения мухи, ее способность стабилизировать движущийся фон восстанавливается. Это указывает на то, что хотя жужжальцы не требуются для обработки движущегося глаза, они вносят в нее свой вклад в зависимости от контекста, когда поведение отделено даже от вращающегося тела. Контекст определяет, будет ли муха использовать свои жужжальцы или зрение в качестве основного источника информации о положении тела / головы.
Необходимость использования жужжальцев в полете была хорошо задокументирована, но мало что известно об их использовании в других формах поведения, таких как ходьба. Некоторые мухи из семейств Muscidae, Anthomyiidae, Calliphoridae, Sarcophagidae, Tachinidae и Micropezidae были задокументированы, чтобы раскачивать крылья во время ходьбы, а также во время полета. Колебания жужжальца аналогичны по амплитуде и частоте во время ходьбы и полета для этих мух, а жужжальца всегда колеблются при ходьбе или в полете. Все остальные семейства двукрылых никогда не раскачивают жужжальца во время ходьбы, но всегда делают это во время полета. Мухи-плоти относятся к тех, которые раскачивают свои жужжальцы во время ходьбы, а также хуже справляются с некоторыми задачами ходьбы, когда их жужжальцы удалены. Напротив, плодовые мушки, которые не раскачивают свои жужжальцы при ходьбе, не проявляют никаких различий в способностях, когда их жужжальцы удалены. Это указывает на то, что сигналы жужжальца имеют поведенческое отношение к тем видам, которые заставляют их колебаться во время ходьбы, и что они помогают этим людям в поведении при ходьбе.