
Войти
| Терминальная полоса | |
|---|---|
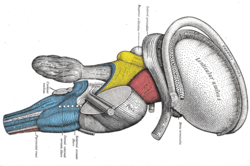 Рассечение ствола головного мозга. Боковой вид. (Терминальная полоска обозначена вверху справа.) Рассечение ствола головного мозга. Боковой вид. (Терминальная полоска обозначена вверху справа.) | |
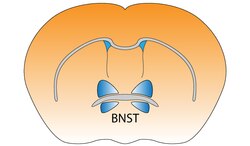 Кровать-ядро терминальной полоски мозга мыши Кровать-ядро терминальной полоски мозга мыши | |
| Подробности | |
| Идентификаторы | |
| латинский | Терминальная полоска |
| NeuroNames | 286 |
| НейроЛекс ID | birnlex_937 |
| TA98 | A14.1.09.275 |
| TA2 | 5592 |
| FMA | 61974 |
| Анатомические термины нейроанатомии [ редактировать в Викиданных ] | |
Терминальная полоска (или терминал полоска ) представляет собой структуру, в головном мозге, состоящий из группы волокон, проходящих вдоль бокового края желудочковой поверхности таламуса. Являясь основным выходным путем миндалины, терминальная полоска проходит от ее центромедиального отдела до вентромедиального ядра гипоталамуса.
Терминальная полоска покрывает верхнюю таламостриатную вену, отмечая линию разделения между таламусом и хвостатым ядром, как видно при крупном разрезе желудочков мозга, если смотреть с верхней стороны.
Терминальная полоска проходит от области межжелудочковых отверстий в височный рог из бокового желудочка, неся волокнами от миндалины к перегородочным ядрам, гипоталамусу и таламус областям головного мозга. Он также переносит волокна, выходящие из этих областей, обратно в миндалину.
Активность ядра ложа терминальной полоски коррелирует с тревогой в ответ на мониторинг угрозы. Считается, что он действует как ретранслятор в системе гипоталамус-гипофиз-надпочечники и регулирует его активность в ответ на острый стресс. Однако реакция на стресс зависит от времени, и BNST не активируется из-за контекстного страха. Это означает, что внезапная пугающая ситуация продолжительностью менее десяти минут не активирует BNST. Также считается, что он способствует поведенческому торможению в ответ на действия незнакомых людей за счет сигналов орбитофронтальной коры. Было показано, что двустороннее нарушение этого пути ослабляет возобновление поведения, связанного с поиском наркотиков, у грызунов.
Это ядро, как известно, проецирует тормозные волокна на боковой гипоталамус и участвует в контроле питания у грызунов. Оптогенетическая активация этого пути ингибирования быстро вызвала у сытых мышей ненасытное пищевое поведение, а оптогенетическое ингибирование этого пути снижает потребление пищи даже у голодных животных.
Центральное подразделение ядра ложа терминальной полоски (BSTc) имеет половой диморфизм. В среднем BSTc в два раза больше у мужчин, чем у женщин, и содержит в два раза больше нейронов соматостатина. В образце шести транс-женщин, подвергнутых посмертной долгосрочной заместительной гормональной терапии (ЗГТ) (от мужчины к женщине), было обнаружено типичное для женщин количество клеток в BSTc, тогда как у транс-мужчины (от женщины к женщине) -мужчина), как было обнаружено, имеет типичный для мужчин номер. Авторы (Jiang-Ning Zhou, Frank PM Kruijver, Dick Swaab ) также обследовали субъектов с гормональными расстройствами и не обнаружили никакой закономерности между этими расстройствами и BSTc, в то время как у одного нелеченного транссексуала из мужчины в женщину было типичное для женщин количество клетки. Они пришли к выводу, что BSTc предоставляет доказательства нейробиологической основы гендерной идентичности, и предположили, что она была определена до рождения.
Было показано, что заместительная гормональная терапия влияет на размер гипоталамуса, даже несмотря на то, что в исследовании пытались сделать это путем включения контрольной группы мужчин и женщин, не являющихся транссексуалами, у которых по разным медицинским причинам произошла гормональная инверсия. Утверждение о нейробиологической основе от рождения позже было поставлено под сомнение, хотя и не опровергнуто, в последующем исследовании той же группы, которое обнаружило, что половой диморфизм BSTc не проявляется до взрослого возраста (примерно 22 года) даже хотя транссексуалы сообщают, что осознают свою гендерную идентичность с детства.
Поскольку нейроны, экспрессирующие соматостатин, как правило, блокируют дендритные входы в постсинаптический нейрон, тем самым подавляя сигналы, проходящие через связанные структуры, считается, что более крупное ядро ложа терминальной полоски, обнаруженное у мужчин (включая трансгендерных мужчин), снижает реакцию испуга у мужчин и может нести ответственность за более высокую частоту конкретных фобий у женщин и возможный источник стереотипа о том, что женщины боятся мышей.
Активность рецептора окситоцина в BNST важна для социального признания у крыс. Как самцы, так и самки крыс, которым вводили микроинъекцию антагониста рецептора окситоцина, имели более низкие оценки социального распознавания, чем крысы, получившие инъекцию носителя, а микроинъекции окситоцина в BNST улучшали социальную память у самцов, но не самок крыс.
Уменьшение размера ядра ложа терминальной полоски наблюдалось у преступников- педофилов мужского пола, в дополнение к уменьшению правой миндалины, гипоталамуса и аномалий в связанных структурах. Авторы предполагают, что детский дефицит BNST и медиальной миндалины может вызывать торможение половой зрелости.
{kind=link}