
Войти
 Просо соматических зародышей
Просо соматических зародышей Соматический эмбриогенез - это искусственный процесс, в котором растение или эмбрион происходит от единственной соматической клетки. Соматические зародыши образуются из растительных клеток, которые в норме не участвуют в развитии зародышей, то есть из обычной растительной ткани. Вокруг соматического зародыша не образуется эндосперм или семенная оболочка.
Клетки, полученные из компетентной исходной ткани, культивируют с образованием недифференцированной массы клеток, называемой каллусом. Регуляторами роста растений в среде для культивирования ткани можно манипулировать, чтобы вызвать образование каллуса, а затем изменить, чтобы побудить эмбрионы формировать каллус. Соотношение различных регуляторов роста растений, необходимых для индукции образования каллуса или зародыша, варьируется в зависимости от типа растения. Соматические зародыши в основном получают in vitro и для лабораторных целей с использованием твердых или жидких питательных сред, которые содержат регуляторы роста растений (PGR). Основными используемыми PGR являются ауксины, но они могут содержать цитокинин в меньшем количестве. Побеги и корни монополярны, в то время как соматические зародыши биполярны, что позволяет им формировать целое растение без культивирования на нескольких типах сред. Соматический эмбриогенез служит моделью для понимания физиологических и биохимических событий, происходящих во время процессов развития растений, а также компонентом биотехнологического прогресса. Первые документы о соматическом эмбриогенезе были получены Steward et al. в 1958 г. и Рейнерт в 1959 г. с суспензионными культурами клеток моркови.
Было описано, что соматический эмбриогенез происходит двумя способами: прямо или косвенно.
происходит, когда эмбрионы запускаются непосредственно из ткани эксплантата, создавая идентичный клон.
происходит, когда эксплантаты продуцируют недифференцированные или частично дифференцированные клетки (часто называемые каллусом), которые затем сохраняются или дифференцируются в ткани растения, такие как лист, стебель или корни. 2,4-дихлорфеноксиуксусная кислота (2,4-D), 6-бензиламинопурин (BAP) и гибберелловая кислота (GA) была использована для разработки непрямые соматические зародыши земляники (Fragaria ananassa)
Регенерация растений посредством соматического эмбриогенеза происходит в пять этапов: инициация эмбриогенных культур, пролиферация эмбриогенных культур, недоношенность соматических зародышей, созревание соматических зародышей и развитие растений на неспецифических средах. Инициирование и пролиферация происходят в среде, богатой ауксином, который индуцирует дифференцировку локализованных меристематических клеток. Обычно используемый ауксин представляет собой 2,4-D. После переноса в среду с низким содержанием ауксина или без него эти клетки могут развиваться в зрелые эмбрионы. Прорастание соматического зародыша может произойти только тогда, когда он достаточно зрел, чтобы иметь функциональные корень и побег верхушки
Факторы и механизмы, контролирующие клеточная дифференцировка у соматических эмбрионов относительно неоднозначна. Было показано, что некоторые соединения, выделяемые культурами тканей растений и обнаруживаемые в питательной среде, необходимы для координации деления клеток и морфологических изменений. Эти соединения были идентифицированы Chung et al. в виде различных полисахаридов, аминокислот, регуляторов роста, витаминов, низкомолекулярных соединений и полипептидов. Было обнаружено несколько сигнальных молекул, которые, как известно, влияют или контролируют образование соматических зародышей, и включают внеклеточные белки, арабиногалактановые белки и липохитоолигосахариды. Температура и освещение также могут влиять на созревание соматического эмбриона.
Применения этого процесса включают: генетически однородный растительный материал; устранение вирусов ; предоставление исходной ткани для генетической трансформации ; создание целых растений из отдельных клеток, называемых протопластами ; разработка технологии синтетических семян.
Разработка процедур соматического эмбриогенеза дала толчок к исследованиям запасных белков семян (SSP) древесных растений для видов деревьев, имеющих коммерческое значение, то есть в основном голосеменных, включая ель белую. В этой области исследований SSP используются в качестве маркеров для определения эмбриогенного потенциала и способности эмбриогенной системы продуцировать соматический эмбрион, биохимически подобный его зиготическому аналогу (Flinn et al. 1991, Beardmore et al. 1997).
Grossnickle et al. (1992) сравнили внутреннюю ель саженцы с приростом во время развития питомника и с помощью программы оценки качества стада непосредственно перед выездом в поле. Высота проростков, диаметр корневой шейки и сухой вес увеличивались у проростков с большей скоростью, чем у зародышей, в течение первой половины первого вегетационного периода, но после этого рост побегов был одинаковым для всех растений. К концу вегетационного периода проростки были на 70% выше, чем зародыши, имели больший диаметр корневой шейки и большую сухую массу побегов. Сухая масса корней у проростков увеличивалась быстрее, чем у зародышей, в начале вегетационного периода
Во время осенней акклиматизации картина увеличения индекса высвобождения покоя и повышения устойчивости к замораживанию была сходной как у проростков, так и у посадок. При осенней акклиматизации способность к росту корней снижалась, а затем увеличивалась, причем у проростков это увеличение было больше.
Оценка качества стада непосредственно перед посадкой показала, что: у закваски было больше эффективности использования воды с уменьшением предрассветного водного потенциала побегов по сравнению с сеянцами; саженцы и зародыши имели одинаковую способность движения воды как при высокой, так и при низкой температуре корней; чистый фотосинтез и проводимость хвои при низких температурах корней были выше у проростков, чем у закваски; и у проростков был больший рост корней, чем у зародышей при температуре корня 22 ° C, но рост корней у всех растений был низким при температуре корня 7,5 ° C.
Рост и выживаемость сеянцев и посадок стироблока внутренней ели 313B после высадки на участке лесовозобновления определяли Grossnickle and Major (1992). Как для проростков, так и для зародышей осмотический потенциал при насыщении (ψ sat) и точка потери тургора (ψ tip) увеличились с минимальных значений -1,82 и -2,22 МПа, соответственно, непосредственно перед к посадке до сезонного максимума -1,09 и -1,21 МПа соответственно при активном удлинении побегов. После этого количество всходов и посадок (ψ sat) и (ψ tip) снизилось до -2,00 и -2,45 МПа соответственно в конце вегетационного периода, что совпало с устойчивым понижение температуры участка и прекращение роста высоты. В целом, проростки и посадки имели одинаковые значения ψ sat и ψ tip в течение вегетационного периода, а также имели аналогичные сдвиги в сезонных моделях максимального модуля упругости, симпатической фракции и относительной содержание воды при точке потери тургора.
Гроссникл и Мейджор (1992) обнаружили, что хвоя однолетних и текущих годов как проростков, так и зародышей имела одинаковое снижение проводимости хвои с увеличением дефицита давления пара. Поверхностные модели отклика нетто-фотосинтеза хвои текущего года (P n), реакции на дефицит давления пара (VPD) и фотосинтетически активную радиацию (PAR) показали, что у эмблингов P n на 15% больше. при VPD менее 3,0 кПа и PAR более 1000 мкмоль мс. Хвоя саженцев и посевов годовалого и текущего года показала схожие модели эффективности использования воды.
Темпы роста побегов у сеянцев и посадок в течение вегетационного периода также были похожи друг на друга. У сеянцев была более крупная система побегов как во время посадки, так и в конце вегетационного периода. У проростков также было более сильное развитие корней, чем у зародышей в течение вегетационного периода, но соотношение корня: побеги для двух типов подвоя было одинаковым в конце вегетационного периода, когда выживаемость сеянцев и заквасок составляла 96% и 99% соответственно.
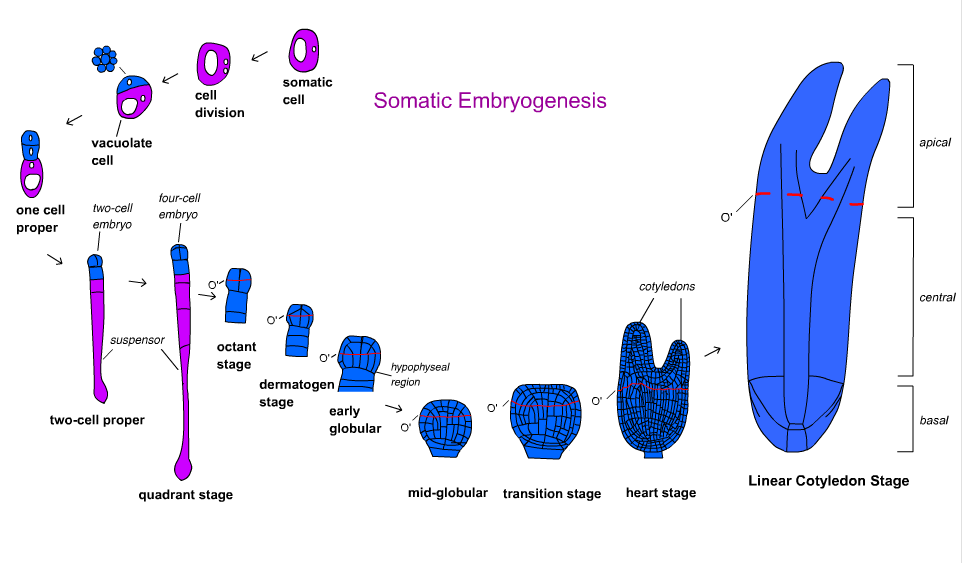
Понимание формирования соматического эмбриона посредством установления морфологических и молекулярных маркеров важно для построения карты судеб. Карта судьбы - это фундамент для дальнейших исследований и экспериментов. Существуют два метода построения карты судьбы: синхронное деление клеток и покадровое отслеживание. Последнее, как правило, работает более стабильно из-за химических веществ, изменяющих клеточный цикл, и центрифугирования, участвующих в синхронном делении клеток.
Развитие зародыша у покрытосеменных растений делится на несколько этапов. Зигота разделена асимметрично, образуя небольшую апикальную клетку и большую базальную клетку. Организационная модель формируется на стадии шаровидного тела, а затем эмбрион переходит в стадию семядоли. Развитие зародыша отличается у однодольных и двудольных. Двудольные проходят через стадию шаровидной, сердцевидной и торпедной формы, в то время как однодольные проходят через стадию шаровидной, щитковой и жесткопиральной.
Многие системы культивирования индуцируют и поддерживают соматический эмбриогенез путем непрерывного воздействия 2,4- дихлорфеноксиуксусная кислота. Абсцизовая кислота, как сообщается, вызывает соматический эмбриогенез у проростков. После формирования каллуса культивирование на среде с низким содержанием ауксина или без гормонов будет способствовать соматическому росту зародыша и формированию корней. У однодольных эмбриогенная способность обычно ограничивается тканями эмбриогенного или меристематического происхождения. Соматические клетки однодольных быстро дифференцируются, а затем теряют митотическую и морфогенную способность. Различия в чувствительности к ауксину при росте эмбриогенного каллуса между разными генотипами одного и того же вида показывают, насколько разнообразными могут быть ответы на ауксин.
Carrot Daucus carota был первым и наиболее изученным видом в отношении путей развития и молекулярных механизмов. Покадровое отслеживание Toonen et al. (1994) показали, что морфология компетентных клеток может варьироваться в зависимости от формы и плотности цитоплазмы. Из эмбриональной суспензии было идентифицировано пять типов клеток: сферические клетки с богатой цитоплазмой, сферические вакуолизированные, овальные вакуолизированные, удлиненные вакуолизированные и клетки неправильной формы. Каждый тип клеток размножается с определенной геометрической симметрией. Они превратились в симметричные, асимметричные кластеры клеток аберрантной формы, которые в конечном итоге сформировали эмбрионы с разной частотой. Это указывает на то, что полярность организованного роста не всегда существует в соматическом эмбриогенезе.
Эмбриональное развитие голосеменных происходит в три фазы. Проэмбриогенез включает все стадии, предшествующие удлинению суспензора. Ранний эмбриогенез включает все стадии после удлинения суспензора, но до развития корневой меристемы. Поздний эмбриогенез включает развитие меристем корня и побега. Покадровая съемка ели обыкновенной Picea abies показала, что ни отдельные богатые цитоплазмой клетки, ни вакуолизированные клетки не развивались в эмбрионы. Проэмбриогенные массы (PEM), промежуточное звено между неорганизованными клетками и эмбрионом, состоящим из богатых цитоплазмой клеток рядом с вакуолизированной клеткой, стимулируются ауксином и цитокинином. Постепенное удаление ауксина и цитокинина и введение абсцизовой кислоты (ABA) позволит сформироваться эмбриону. Использование соматического эмбриогенеза было рассмотрено для массового производства вегетативно размножаемых клонов хвойных деревьев и криоконсервации зародышевой плазмы. Однако использование этой технологии для лесовосстановления и выращивания хвойных деревьев находится в зачаточном состоянии.
{kind=link}