
Войти
Контралатеральный организация переднего мозга (латинское : contra «против»; latus «боковая», латеральная «боковая») - это свойство, согласно которому полушария головного мозга и таламус представляют в основном контралатеральную сторону тела. Следовательно, левая часть переднего мозга в основном представляет собой правую сторону тела, а правая часть мозга представляет в основном левую часть тела. Контралатеральная организация задействует как исполнительные, так и сенсорные функции (например, левостороннее поражение мозга может вызвать правостороннюю гемиплегию ). Контралатеральная организация присутствует у всех позвоночных, но не у беспозвоночных.
Согласно современной теории, передний мозг скручен вокруг длинной оси тела, так что не только левый и правый стороны, но также чередуются дорсальная и вентральная стороны (см. ниже).
 Некоторые афферентные перекресты.
Некоторые афферентные перекресты.  пирамидные перекресты.
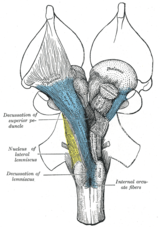
пирамидные перекресты. Анатомически контралатеральная организация проявляется в основных дискуссиях (латинское: латинское обозначение для десять, 'дека', является заглавной 'X') и хиазмой (после греческого заглавной буква 'Χ', чи ). Перекрест означает пересечение пучков аксональных волокон внутри центральной нервной системы. В результате таких перекрестков эфферентные связи головного мозга с базальными ганглиями, мозжечком и позвоночником пересекаются, и афферентные связи от позвоночника, мозжечка и моста с таламусом пересекаются. В результате моторные, соматосенсорные, слуховые и зрительные первичные области в переднем мозге представляют преимущественно контралатеральную сторону тела.
Два из черепных нервов демонстрируют перекрестье: перекрест зрительного тракта (черепной нерв II), который берет свое начало из глаз и вставляет на оптическом слое среднего мозга и на блокированном нерве (нерв IV), который берет начало в брюшной части среднего мозга и иннервирует одну из шести мышц, которые вращают глаз (верхняя косая мышца ).
Хотя передний мозг всех позвоночных имеет контралатеральную организацию, эта контралатеральность ни в коем случае не является полной. Стоит упомянуть некоторые из этих исключений:
Согласно современным представлениям, контралатеральная организация возникает из-за осевого скручивания (см. ниже), это единственная теория, которая была протестирована независимо. Ранее был опубликован ряд альтернативных предложений, наиболее популярным из которых является теория визуальных карт . научно-популярное видео объясняет эти теории вкратце.
 Схема теории визуальных карт Кахала. O = зрительный перекрест; C = зрительная (и двигательная) кора; M, S = переходные пути; R, G: Сенсорные нервы, двигательные ганглии.
Схема теории визуальных карт Кахала. O = зрительный перекрест; C = зрительная (и двигательная) кора; M, S = переходные пути; R, G: Сенсорные нервы, двигательные ганглии. Теория визуальных карт была опубликована известным нейробиологом и пионером Сантьяго Рамон-и-Кахал (1898), см. Также и английские резюме. Согласно этой теории, функция перекреста зрительных нервов состоит в восстановлении изображения поля сетчатки на зрительной коре. Зрачок в глазах позвоночных инвертирует изображение на сетчатке, так что визуальная периферия проецируется на медиальную сторону сетчатки. Благодаря хиазматическому пересечению зрительная периферия снова оказывается снаружи, если предположить, что карта сетчатки точно сохраняется по всему зрительному тракту.
У теории есть ряд слабых мест. Например, зрительные тракты растягиваются по спирали от таламического LGN к зрительной коре (см. Рисунок; этот путь известен как оптическое излучение ). В результате карта сетчатки показывает визуальную периферию на медиальной стороне. Однако центральным моментом теории было именно получение точной визуальной карты с проекцией медиального поля на медиальные стороны зрительной коры.
Две теории скручивания были предложены независимо: Гипотеза осевого скручивания де Марка Люссане и Яна Оссе и гипотеза соматического скручивания Марселя Кинсборна.
 Схема эволюционного скручивания согласно гипотеза осевого скручивания. А, В: Ранний зародыш поворачивается на левую сторону; B, C: Симметрия сохраняется за счет дальнейшего поворота влево в передней части головы и компенсирующего поворота вправо в остальной части тела. D, E: Рост зрительного тракта, ведущий к перекресту зрительных нервов. Цвета относятся к раннему эмбриону: красный = правая сторона, синий = левая сторона, черный = дорсальный, белый = вентральный
Схема эволюционного скручивания согласно гипотеза осевого скручивания. А, В: Ранний зародыш поворачивается на левую сторону; B, C: Симметрия сохраняется за счет дальнейшего поворота влево в передней части головы и компенсирующего поворота вправо в остальной части тела. D, E: Рост зрительного тракта, ведущий к перекресту зрительных нервов. Цвета относятся к раннему эмбриону: красный = правая сторона, синий = левая сторона, черный = дорсальный, белый = вентральный Гипотеза осевого скручивания была разработана, чтобы объяснить, каким образом структура контралатеральной организации, перекрестов и хиазмов развивается, и почему этот образец эволюционно устойчив. Эволюционная стабильность действительно замечательна, учитывая, что на протяжении 500 миллионов лет эволюции позвоночных не было известных исключений. Согласно теории, контралатеральная организация развивается следующим образом. Ранний эмбрион поворачивают на левую сторону, так что его левая сторона обращена к желтку, а правая - к желтку. Эта асимметричная ориентация компенсируется асимметричным ростом для восстановления поверхностной двусторонней симметрии. Передняя часть головы поворачивается влево, как показано на схеме. Передний мозг не является поверхностной структурой, но он настолько тесно связан с поверхностными структурами тела, что поворачивается вместе с передней частью головы. Эти структуры позже сформируют глаза, ноздри и рот.
Тело за головой компенсирует асимметричную ориентацию тела в противоположном направлении поворотом вправо (см. Схему). Из-за этих противоположно направленных компенсаций передней части головы и остального тела животное искривляется.
Зрительный тракт растет от сетчатки к оптической оболочке, поскольку дорсальный и вентральный перевернуты в передней области головы, тракты сначала растут к вентральной стороне, чтобы встретиться по средней линии, образуя хиазму. Поскольку покров зрительного нерва лежит на дорсальной части среднего мозга, каждый тракт затем продолжается дорсально до контралатеральной оболочки зрительного нерва.
Сердце и кишечник - это внутренние органы, не имеющие прочной интеграции с внешними структурами тела, поэтому эволюционное давление не заставляет их также вращаться. Скорее, эти органы сохраняют свою изначальную асимметричную ориентацию в теле.
Гипотеза осевого скручивания предсказывает, что небольшая асимметрия лица, мозга и туловища (в противоположном направлении) сохраняется и во взрослой жизни, и это было подтверждено экспериментально.
Идея соматического поворота была вдохновлена гипотезой дорсовентральной инверсии ; идея была разработана Марселем Кинсборном.
Согласно гипотезе дорсовентральной инверсии, предковый дейтеростом перевернулся на спину. В результате у позвоночных имеется дорсальная нервная система, а у протостомов вентральная. Согласно гипотезе соматического поворота, не все животное перевернулось на спину, а только «соматическая» часть, то есть все, что находится за глазами, ртом и ноздрями, включая передний мозг.
Эти три теории тесно связаны. Гипотеза соматического поворота была предложена как усовершенствование гипотезы инверсии и, таким образом, имеет гораздо более широкую объяснительную силу, чем ее предшественник. Это объясняет не только инверсию тела, но и противоположный передний мозг. Гипотеза осевого скручивания была определена независимо от двух других. Помимо перевернутого тела и контралатерального переднего мозга, это объясняет, почему сердце и кишечник асимметричны. Более того, это единственное из трех, подтвержденное данными эмбриологического роста.
Замечательным свойством контралатеральной организации является то, что она присутствует у каждого позвоночного. Даже самые далекие клады agnathans обладают зрительным перекрестом, и даже отпечатки черепа ранних позвоночных из ордовика демонстрируют наличие оптического перекреста: эта идея была разработана Кинсборном. Есть молекулярные доказательства гипотезы инверсии почти для всех групп дейтеростом. Однако неизвестно, какое именно давление отбора вызвало инверсию. Скручивание и асимметричное развитие хорошо известны у других дейтеростомий, таких как Hemichordata, Echinodermata, Cephalochordata и Tunicata. В этих кладах также часто встречается поворот на бок или вверх ногами.
При голопрозэнцефалии полушария головного мозга или его часть выравниваются не с левой и правой стороны, а с фронтальной и затылочной стороны черепа., и обычно остается очень маленьким. Согласно гипотезе осевого скручивания, это представляет собой крайний случай яковлевского крутящего момента и может возникнуть, когда головной мозг не поворачивается во время ранней эмбриологии.
Близнецы Cephalopagus или janiceps - это сиамские близнецы, которые рождаются с двумя лицами, по одному с обеих сторон головы. У этих близнецов два мозга и два спинных мозга, но они расположены с левой и с правой стороны тела. Согласно гипотезе осевого скручивания, две нервные системы не могли поворачиваться из-за сложной конфигурации тела и поэтому оставались по обе стороны.