Трофическая когерентность - это свойство ориентированных графов (или направленных сетей ). Он основан на концепции трофических уровней, используемых в основном в ecology, но которые могут быть определены для направленных сетей в целом и обеспечивают меру иерархической структуры между узлами. Трофическая согласованность - это тенденция узлов попадать на четко определенные трофические уровни. Это было связано с несколькими структурными и динамическими свойствами направленных сетей, включая преобладание циклов и сетевых мотивов, экологической стабильности, интервальности и процессов распространения, таких как эпидемии и нейронные лавины.
Содержание
- 1 Определение
- 2 Трофическая когерентность по природе
- 3 Трофические уровни и функция узла
- 4 Создание трофически когерентных сетей
- 5 Ссылки
- 6 Внешние ссылки
Определение
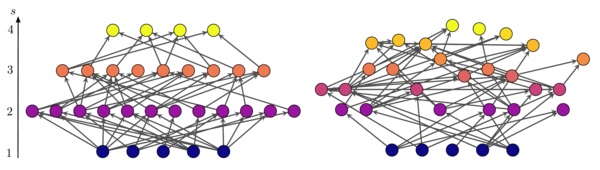
Две направленные сети, созданные с помощью «обобщенной модели предпочтительного преследования». Положение узлов на вертикальной оси представляет их трофический уровень. Сеть слева максимально когерентна (q = 0), а справа - более некогерентна (q = 0,49).
Рассмотрим направленную сеть, определяемую  матрица смежности
матрица смежности  . Каждому узлу
. Каждому узлу  может быть присвоен трофический уровень
может быть присвоен трофический уровень  согласно
согласно

где  - это в градусах, а узлы с
- это в градусах, а узлы с  (базальные узлы) условно имеют
(базальные узлы) условно имеют  . С каждым ребром связано трофическое различие, определяемое как
. С каждым ребром связано трофическое различие, определяемое как  . Трофическая когерентность сети - это мера того, насколько сильно распределение трофических расстояний,
. Трофическая когерентность сети - это мера того, насколько сильно распределение трофических расстояний,  , находится около своего среднего значения, которое всегда
, находится около своего среднего значения, которое всегда  . Это может быть зафиксировано параметром некогерентности
. Это может быть зафиксировано параметром некогерентности  , равным стандартному отклонению :
, равным стандартному отклонению :

где  - количество ребер в сети.
- количество ребер в сети.
На рисунке показаны две сети, различающиеся трофической связностью. Положение узлов на вертикальной оси соответствует их трофическому уровню. В сети слева узлы попадают на отдельные (целые) трофические уровни, поэтому сеть максимально когерентна  . На рисунке справа многие из узлов имеют фракционные трофические уровни, а сеть более непоследовательна
. На рисунке справа многие из узлов имеют фракционные трофические уровни, а сеть более непоследовательна  .
.
Трофическая согласованность по природе
Степень трофической когерентности (или некогерентности) эмпирических сетей может быть исследована путем сравнения с нулевой моделью. Это обеспечивается базовым ансамблем, который включает сети, в которых все небазальные узлы имеют одинаковую долю базальных узлов для своих соседей. Ожидаемые значения в этом ансамбле сходятся со значениями широко используемого конфигурационного ансамбля в пределе  ,
,  (с
(с  и
и  количеством узлов и ребер) и может численно показать, что это хорошее приближение для конечных случайных сетей. Базовое ожидание ансамбля для параметра некогерентности составляет
количеством узлов и ребер) и может численно показать, что это хорошее приближение для конечных случайных сетей. Базовое ожидание ансамбля для параметра некогерентности составляет

где  - количество ребер, соединенных с базальными узлами. Отношение
- количество ребер, соединенных с базальными узлами. Отношение  , измеренное в эмпирических сетях, показывает, являются ли они более или менее согласованными, чем случайное ожидание. Например, Джонсон и Джонс обнаружили в наборе сетей, что пищевые сети значительно когерентны
, измеренное в эмпирических сетях, показывает, являются ли они более или менее согласованными, чем случайное ожидание. Например, Джонсон и Джонс обнаружили в наборе сетей, что пищевые сети значительно когерентны  , метаболические сети значительно некогерентны
, метаболические сети значительно некогерентны  и сети регуляции генов близки к случайному ожиданию
и сети регуляции генов близки к случайному ожиданию  .
.
Трофические уровни и функция узла
Пока еще мало понимания механизмов, которые могут привести к тому, что определенные виды сетей станут значительно связными или несогласованными. Однако в системах, которые представляют корреляции между трофическим уровнем и другими характеристиками узлов, процессы, которые имеют тенденцию способствовать созданию границ между узлами с определенными характеристиками, могут вызывать согласованность или несогласованность. В случае пищевых сетей хищники, как правило, специализируются на потреблении добычи с определенными биологическими свойствами (такими как размер, скорость или поведение), которые коррелируют с их диетой и, следовательно, с трофическим уровнем. Это было предложено как причина согласованности трофической сети. Однако модели трофической сети, основанные на оси ниши, не воспроизводят реалистичную трофическую когерентность, что может означать, что либо этого объяснения недостаточно, либо необходимо учитывать несколько параметров ниши.

Сеть сцепленных слова доктора Сьюза из книги «Зеленые яйца и ветчина». Если слово 1 появляется непосредственно перед словом 2 хотя бы в одном предложении текста, направленный край (стрелка) помещается от слова 1 к слову 2. Высота каждого слова пропорциональна его трофическому уровню. Цвета указывают на синтаксическую функцию; от низшего к высшему средний трофический уровень: существительные (синий), предлоги и союзы (голубой), определители (розовый), наречия (желтый), местоимения (зеленый), глаголы (красный) и прилагательные (фиолетовый). Когда слово имеет более одной функции, используется одна, наиболее часто встречающаяся в тексте.
Связь между трофическим уровнем и функцией узла можно увидеть в сетях, отличных от пищевых сетей. На рисунке показана словесная сеть смежности, полученная из книги Green Eggs and Ham автора Dr Seuss. Высота узлов представляет их трофические уровни (в соответствии с направлением ребер, которое противоположно тому, что указано стрелками, которые указывают порядок, в котором слова объединяются в предложения). Синтаксическая функция слов также отображается цветом узла. Существует четкая взаимосвязь между синтаксической функцией и трофическим уровнем: средний трофический уровень нарицательных существительных (синий) равен  , тогда как у глаголов (красный) это
, тогда как у глаголов (красный) это  . Этот пример показывает, как тропическая когерентность или несогласованность может возникать из-за функции узла, а также то, что трофическая структура сетей обеспечивает средства идентификации функции узла в определенных системах.
. Этот пример показывает, как тропическая когерентность или несогласованность может возникать из-за функции узла, а также то, что трофическая структура сетей обеспечивает средства идентификации функции узла в определенных системах.
Создание трофически когерентных сетей
Существуют различные способы создания направленных сетей с заданной трофической когерентностью, и все они основаны на постепенном введении новых ребер в систему таким образом, чтобы вероятность каждого нового кандидата Принятие края зависит от ожидаемой трофической разницы.
Модель предпочтительной добычи - это развивающаяся сетевая модель, аналогичная модели Барбаси-Альберта предпочтительной привязанности, но вдохновленная экосистемой, которая растет за счет иммиграции новых видов. Первый начинается с  базальных узлов и продолжается введением новых узлов до . Каждому новому узлу назначается первый ближайший сосед
базальных узлов и продолжается введением новых узлов до . Каждому новому узлу назначается первый ближайший сосед  (вид добычи в контексте пищевой сети) и новый край размещается от до . Новому узлу присваивается временный трофический уровень
(вид добычи в контексте пищевой сети) и новый край размещается от до . Новому узлу присваивается временный трофический уровень  . Затем для из числа участников сети в соответствии с их трофическими уровнями. В частности, для нового кандидата в соседние объекты
. Затем для из числа участников сети в соответствии с их трофическими уровнями. В частности, для нового кандидата в соседние объекты  вероятность быть выбранным является функцией
вероятность быть выбранным является функцией  . Джонсон и другие используют
. Джонсон и другие используют

где  - параметр, который настраивает трофическую когерентность: для
- параметр, который настраивает трофическую когерентность: для  генерируются максимально когерентные сети, а монотонно увеличивается с для
генерируются максимально когерентные сети, а монотонно увеличивается с для  . Выбор
. Выбор  является произвольным. Один из возможных вариантов - установить значение
является произвольным. Один из возможных вариантов - установить значение  , где
, где  - количество узлов уже в сети на момент прибытия , а
- количество узлов уже в сети на момент прибытия , а  - случайная величина получено из бета-распределения с параметрами
- случайная величина получено из бета-распределения с параметрами  и
и

( желаемое количество ребер). Таким образом, обобщенная каскадная модель восстанавливается в пределе
желаемое количество ребер). Таким образом, обобщенная каскадная модель восстанавливается в пределе  , а распределения степеней такие же, как в нишевой модели и обобщенной нишевой модели. Этот алгоритм, как описано выше, генерирует сети без циклов (за исключением самоциклов, если новый узел сам считается среди своих соседей-кандидатов ). Чтобы циклы любой длины были возможны, можно рассмотреть новые ребра-кандидаты, в которых новый узел является внутренним соседом, а также те, в которых он будет чужим соседом. Вероятность принятия этих ребер,
, а распределения степеней такие же, как в нишевой модели и обобщенной нишевой модели. Этот алгоритм, как описано выше, генерирует сети без циклов (за исключением самоциклов, если новый узел сам считается среди своих соседей-кандидатов ). Чтобы циклы любой длины были возможны, можно рассмотреть новые ребра-кандидаты, в которых новый узел является внутренним соседом, а также те, в которых он будет чужим соседом. Вероятность принятия этих ребер,  , затем зависит от
, затем зависит от  .
.
Обобщенная модель предпочтительного преследования аналогична описанной выше, но имеет определенные преимущества. В частности, он более поддается анализу, и можно создавать сети с точным числом ребер . Сеть начинается с базальных узлов, а затем еще  добавляются новые узлы в следующих путь. Когда каждый входит в систему, ему случайным образом назначается один ближайший сосед из числа уже имеющихся. Тогда каждый узел имеет целочисленный временный трофический уровень
добавляются новые узлы в следующих путь. Когда каждый входит в систему, ему случайным образом назначается один ближайший сосед из числа уже имеющихся. Тогда каждый узел имеет целочисленный временный трофический уровень  . Остальные ребра
. Остальные ребра  вводятся следующим образом. Каждая пара узлов
вводятся следующим образом. Каждая пара узлов  имеет два связанных временных трофических расстояния,
имеет два связанных временных трофических расстояния,  и
и  . Каждое из этих возможных ребер принимается с вероятностью, которая зависит от этого временного расстояния. Клэйз и Джонсон используют
. Каждое из этих возможных ребер принимается с вероятностью, которая зависит от этого временного расстояния. Клэйз и Джонсон используют

, потому что они считают, что распределение трофических расстояний в нескольких типах сетей приблизительно нормальное, и этот выбор приводит к диапазону параметра , в котором  . После того, как все ребра введены, необходимо пересчитать трофические уровни всех узлов, поскольку они будут отличаться от первоначально назначенных временных, если только
. После того, как все ребра введены, необходимо пересчитать трофические уровни всех узлов, поскольку они будут отличаться от первоначально назначенных временных, если только  . Как и в случае с моделью предпочтительного преследования, средний параметр некогерентности результирующих сетей является монотонно возрастающей функцией от для
. Как и в случае с моделью предпочтительного преследования, средний параметр некогерентности результирующих сетей является монотонно возрастающей функцией от для  . На рисунке выше показаны две сети с разной трофической когерентностью, созданные с помощью этого алгоритма.
. На рисунке выше показаны две сети с разной трофической когерентностью, созданные с помощью этого алгоритма.
Ссылки
Внешние ссылки

 Две направленные сети, созданные с помощью «обобщенной модели предпочтительного преследования». Положение узлов на вертикальной оси представляет их трофический уровень. Сеть слева максимально когерентна (q = 0), а справа - более некогерентна (q = 0,49).
Две направленные сети, созданные с помощью «обобщенной модели предпочтительного преследования». Положение узлов на вертикальной оси представляет их трофический уровень. Сеть слева максимально когерентна (q = 0), а справа - более некогерентна (q = 0,49).  Сеть сцепленных слова доктора Сьюза из книги «Зеленые яйца и ветчина». Если слово 1 появляется непосредственно перед словом 2 хотя бы в одном предложении текста, направленный край (стрелка) помещается от слова 1 к слову 2. Высота каждого слова пропорциональна его трофическому уровню. Цвета указывают на синтаксическую функцию; от низшего к высшему средний трофический уровень: существительные (синий), предлоги и союзы (голубой), определители (розовый), наречия (желтый), местоимения (зеленый), глаголы (красный) и прилагательные (фиолетовый). Когда слово имеет более одной функции, используется одна, наиболее часто встречающаяся в тексте.
Сеть сцепленных слова доктора Сьюза из книги «Зеленые яйца и ветчина». Если слово 1 появляется непосредственно перед словом 2 хотя бы в одном предложении текста, направленный край (стрелка) помещается от слова 1 к слову 2. Высота каждого слова пропорциональна его трофическому уровню. Цвета указывают на синтаксическую функцию; от низшего к высшему средний трофический уровень: существительные (синий), предлоги и союзы (голубой), определители (розовый), наречия (желтый), местоимения (зеленый), глаголы (красный) и прилагательные (фиолетовый). Когда слово имеет более одной функции, используется одна, наиболее часто встречающаяся в тексте.