
Войти
| Creodonta. Временной диапазон: от раннего палеоцена до позднего Миоцен, 63.3–11.1 Ma PreꞒ Ꞓ O S D C P T J K Pg N | |
|---|---|
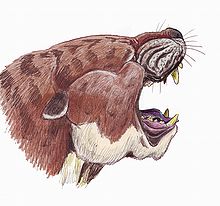 | |
| Sarkastodon | |
| Научная классификация | |
| Царство: | Animalia |
| Тип: | Chordata |
| Класс: | Mammalia |
| Clade: | Ferae |
| Порядок: | †Creodonta. (Cope, 1875) |
| Семьи | |
Creodonta является вымершие, возможно, полифилетические отряд плотоядных млекопитающих, которые жили с палеоцена до миоцена эпохи. Поскольку они оба обладают хищными зубами, когда-то считалось, что креодонты и хищники имели общего предка, но учитывая, что в состав хищников вовлечены разные зубы (как у креодонтов, так и у хищников) и между основными группами креодонтов), это, по-видимому, случай эволюционной конвергенции. Плотоядные животные также известны у других кладов млекопитающих, например, у вымерших летучих мышей Necromantis.
Исторически считалось, что в отряд входят два разных семейства: Oxyaenidae и Hyaenodontidae. Оба они могут представлять отдельные отряды ископаемых млекопитающих, связанных с карнивораморфами, или происходить от более базальных таксонов.
Креодонты имели обширный ареал, как географически, так и временно. Они известны от позднего палеоцена до позднего олигоцена в Северной Америке, от раннего эоцена до позднего олигоцена в Европе, от раннего эоцена до позднего миоцена в Азии и от палеоцена до позднего миоцена в Африке.
Креодонты были первыми крупными, очевидно хищными млекопитающими, получившими излучение оксиенид в позднем палеоцене. Во время палеогена они были самой многочисленной формой наземных хищников в Старом Свете. В олигоцене Африке они были доминирующей группой хищников. Они конкурировали с мезонихидами и энтелодонтами и в конечном итоге пережили их к началу олигоцена и к середине миоцена соответственно, но уступили позиции плотоядные. Последний род вымер 11,1 миллионов лет назад, и теперь плотоядные животные занимают свои экологические ниши.
 Гора оксианидов Патриофелис из Американский музей естественной истории.
Гора оксианидов Патриофелис из Американский музей естественной истории. «Креодонта» была придумана Эдвардом Дринкером в 1875 году. Коуп включил оксианидов и виверравидов Didymictis, но не включил гиенодонтиды. В 1880 г. он расширил этот термин, включив в него Miacidae, Arctocyonidae, (теперь), Oxyaenidae и Mesonychidae. Коп изначально поместил креодонтов в насекомоядных. Однако в 1884 году он рассматривал их как базальную группу, из которой произошли как плотоядные, так и насекомоядные. Hyaenodontidae не входили в число креодонтов до 1909 года. Со временем различные группы были удалены, и к 1969 году они содержали: как и сегодня, только оксиениды и гиенодонтиды.
 Череп оксианид Machaeroides eothen.
Череп оксианид Machaeroides eothen. Одно из взглядов на положение группы состоит в том, что Creodonta и Carnivora являются сестринскими таксонами (в пределах надотряда Ferae). Другие утверждали, что насекомоядные более тесно связаны с плотоядными, а креодонты, следовательно, являются базальными эвтерианами. Другие предположили, что креодонта не может быть монофилетической. Полли утверждал, что единственная доступная синапоморфия между оксиенидами и гиенодонтидами - это большая метастилярная пластинка на первом моляре (M1), но он полагает, что эта особенность является общей для всех базальных эутерий. Отделение Oxyaenidae от Hyaenodontidae также согласуется с биогеографическими данными, поскольку первые оксианиды известны из раннего палеоцена Северной Америки, а первые гиенодонтиды относятся к очень позднему палеоцену Северной Африки.
 Боковой (A) и дорсальный (B) виды на череп гиенодонтида Apterodon macrognathus Генри Фэйрфилд Осборн.
Боковой (A) и дорсальный (B) виды на череп гиенодонтида Apterodon macrognathus Генри Фэйрфилд Осборн.Эту компоновку усложняет предварительное одобрение Ганнеллом возведения третьего семейства, Limnocyonidae. В эту группу входят таксоны, которые когда-то считались оксиенидами, такие как Limnocyon и Prolimnocyon. Вортман даже построил подсемейство Limnocyoninae в оксиенидах. Ван Вален гнездится в том же подсемействе (в том числе) внутри Hyaenodontidae. Ганнелл не знает, входит ли Limnocyonidae в группу Hyaenodontidae (хотя и является сестринской по отношению к остальным гиенодонтидам) или полностью отдельно.
 Sinopa окаменелости: (1) правые верхние щечные зубы, P2-M2; (2) Левая ветвь нижней челюсти (p2-m2); (3) Правая ветвь нижней челюсти (c-m2).
Sinopa окаменелости: (1) правые верхние щечные зубы, P2-M2; (2) Левая ветвь нижней челюсти (p2-m2); (3) Правая ветвь нижней челюсти (c-m2). Согласно Ганнеллу, отличительными чертами оксиенид являются: Небольшая черепная коробка в нижней части черепа. затылочный бугор широкий в основании и сужается дорсально (для придания ему треугольной формы). слезная кость образует полукруглое расширение на лице. На нижней челюсти тяжелый симфиз. M1 и m2 образуют хищники, а M3 / m3 отсутствуют. manus и pes - стопоходящие или субплантиградные. малоберцовая кость сочленяется с пяточной костью, а астрагал сочленяется с кубовидной костью. фаланги сжаты и имеют трещины на кончике.
Аналогичным образом, список определяющих черт гиенодонтид Ганнелла включает: Длинный узкий череп с узким базикранием и высоким узкий затылок. лобные кости вогнуты между орбитальными областями. M2 и m3 образуют хищники. M3 присутствует у большинства видов, а m3 присутствует всегда. Manus и pes варьируются от стопоходящих до пальцевидных. Малоберцовая кость сочленяется с пяточной костью, а астрагалово-кубовидное сочленение сокращено или отсутствует. Концевые фаланги сдавлены и имеют трещины на кончике.
Лимноциониды имели следующие особенности, согласно Ганнеллу: M3 / m3 были уменьшены или отсутствовали, остальные зубы не были восстановлены. Рострум был удлиненным. Сами животные были от маленьких до средних.
В последнее время «креодонта» считалась полифилетическим сообществом млекопитающих, а не естественной группой, а оксиениды считались базальными лауразиатерами. Однако филогенетический анализ палеоценовых млекопитающих, опубликованный в 2015 году, подтвердил монофилию креодонтов и поместил эту группу в родственников панголинов.
 Сравнение хищных зубов волка и типичных гиенодонтид и оксиаенид
Сравнение хищных зубов волка и типичных гиенодонтид и оксиаенид Среди примитивных креодонтов зубная формула - 3.1.4.33.1.4.3, но более поздние формы часто имели уменьшенное количество резцов, премоляров и / или коренных зубов. Клыки всегда большие и заостренные. Боковые резцы большие, а средние резцы обычно маленькие. Премоляры примитивны, с одним первичным бугорком и различными вторичными бугорками.
Креодонты имеют две или три пары хищных зубов, но только одна пара выполняет режущую функцию (либо M1 / m2, либо M2 / м3). Такое расположение отличается от современных хищников, которые используют P4 и m1 для хищников, и это предполагает отдельную историю эволюции и различие на уровне порядка.
В двух основных группах креодонтов были задействованы разные коренные зубы. У Oxyaenidae хищники образуют M1 и m2. Среди гиенодонтид это М2 и м3. В отличие от большинства современных хищников, у которых хищники являются единственными режущими зубами, другие коренные зубы креодонта выполняют второстепенные режущие функции. Разница в том, какие зубы образуют хищные зубы, является основным аргументом в пользу полифилии Креодонты.
 Верхний вид черепа лимноционида (?) Limnocyon verus.
Верхний вид черепа лимноционида (?) Limnocyon verus.У креодонтов были длинные узкие черепа с маленьким мозгом. Череп значительно сужался за глазами, давая отчетливые спланхнокраниальные и нейрокраниальные сегменты черепа. У них были большие сагиттальные гребни и обычно широкие сосцевидные отростки (которые, вероятно, были производными чертами для группы). Многие креодонты имели пропорционально большие головы. У примитивных форм слуховые буллы не окостенели. В целом височные ямки были очень широкими.
 Верховой скелет гиенодонтида Sinopa rapax из бассейна Бриджер.
Верховой скелет гиенодонтида Sinopa rapax из бассейна Бриджер. Креодонты имели обобщенные посткраниальные скелеты.. Их конечности были мезаксоническими (с осью стопы на середине их пяти пальцев). Их метод передвижения варьировался от стопоходящего до пальцевого. Концевые фаланги представляли собой сросшиеся когти.
 Боковой контур и вид спереди черепа Sarkastodon mongoliensis.
Боковой контур и вид спереди черепа Sarkastodon mongoliensis.Креодонты были размером от маленького кота до 800 кг Саркастодон. Однако более крупные животные были известны только в конце палеоцена с излучением оксиенид, таких как размером с пуму и, вероятно, сокрушающий кости падальщик.
Некоторые креодонты (Arfia, Prolimnocyon и Palaeonictis)), похоже, испытали явление карликования во время палеоцен-эоценового теплового максимума, наблюдаемого у других родов млекопитающих. Предлагаемое объяснение этого явления состоит в том, что повышенный уровень углекислого газа в атмосфере напрямую повлиял на плотоядных из-за повышения температуры и засушливости, а также косвенно повлиял на них, уменьшив размер их травоядной жертвы за счет того же давления отбора.
Самый крупный североамериканский креодонт - Patriofelis. Образец P. ferox, собранный в бассейне Бриджер на юге Вайоминга, был размером с взрослого черного медведя с головой почти как взрослый самец льва.
Во время Среднеазиатской экспедиции 1930 г. В Американском музее естественной истории был собран крупнейший из когда-либо обнаруженных креодонтов: Sarkastodon mongoliensis. Его размеры были на 50% больше, чем у Patriofelis, с которым он имел много общего. Было подсчитано, что Саркастодон в два раза превышал массу тела самого большого американского льва.
 Верхние зубы креодонтов из Среднеэоценового бассейна Бриджер, Вайоминг
Верхние зубы креодонтов из Среднеэоценового бассейна Бриджер, Вайоминг Ранние креодонты (как оксиениды, так и гиенодонтиды) имели трибосфеновые коренные зубы, общие для базальных терианцев. У малых форм были довольно сильные постметакристо-метастзвездные гребни, что позволяет предположить, что они, вероятно, были оппортунистическими кормушками, питавшимися такими вещами, как яйца, птицы, мелкие млекопитающие, насекомые и, возможно, растительное вещество, возможно, как современные виверриды. Более крупные формы имели большую режущую способность, и производительность со временем увеличивалась., одно из самых распространенных плотоядных млекопитающих в раннем эоцене Северной Америки, в течение раннего эоцена развило более открытый тригонид на M3, увеличив стригущую способность хищников. Сходное развитие можно увидеть, сравнив и с более мелкими, более обобщенными кормушками среди креодонтов.
 Патриофелис
Патриофелис Креодонты традиционно считались предками хищников, но теперь считаются таковыми. более близок к панголинам. Оксиениды впервые известны из палеоцена Северной Америки, тогда как гиенодонты происходят из палеоцена Африки.
Они разделяют с плотоядными и многими другими кладами хищных млекопитающих хищные ножницы, модификацию зубов, которые эволюционировали, чтобы разрезать мясо, как ножницы, и дали обоим приказ орудиям доминировать в нише - адаптация, также наблюдаемая у других кладов хищных млекопитающих. Их происхождение может быть как минимум еще в конце мелового периода, в зависимости от методов генетической калибровки плаценты, хотя до кайнозоя они не излучали много. Креодонты были доминирующими хищными млекопитающими с 55 до 35 миллионов лет назад, пик разнообразия и распространенности пришелся на эоцен. К середине олигоцена креодонты в конечном итоге вытеснили мезонихидов полностью в Северной Америке и Евразии, претерпев диверсификацию в Африка, в свою очередь, соревновалась со своими родственниками-хищниками. Последний род, Dissopsalis, вымер около 11,1 миллионов лет назад.
Креодонты располагались в Северной Америке, Евразии и Африке по форме, напоминающей формы некоторых современных хищники. Среди них был саркастодон, один из крупнейших наземных хищников всех времен, весивший примерно 800 кг. Их господство над ранними хищниками, известными как миациды, начало ослабевать после 35 миллионов лет назад. Креодонты дожили до 8 миллионов лет назад; последняя форма, Dissopsalis, вымерла в Пакистане. Медведи, кошки, куньи, гиены, собаки и другие хищники теперь занимают бывшие ниши креодонтов.
 Череп гиенодона
Череп гиенодона Точно неизвестно, почему креодонты были заменены хищниками. Это может быть из-за их меньшего размера мозга и их передвижения, которое было несколько менее энергоэффективным (особенно во время бега). Их структура конечностей ограничивала движение ног в вертикальной плоскости, как у лошадей; они не могли повернуть запястья и предплечья внутрь, чтобы споткнуться, порезать или схватить добычу, как современные хищники. Креодонты должны были полностью полагаться на свои челюсти, чтобы поймать добычу, что может быть причиной того, что креодонты обычно имели больший размер головы по сравнению с их телом, чем плотоядные животные того же роста. Пояснично-крестцовый отдел креодонта не был приспособлен для бега так эффективно, как у Carnivora. Расположение зубов тоже было несколько иное. У миацидов (как и у современных Carnivora) последний верхний премоляр и первый нижний коренной зуб являются хищными, позволяя скрежетать зубами на некоторое время. питаются немясной пищей (Canidae - самый близкий современный аналог зубного ряда миацид). У креодонтов хищники были еще дальше - либо первые верхние и вторые нижние коренные зубы, либо вторые верхние и третьи нижние коренные зубы. Это заставляло их есть почти исключительно мясо. Эти ограничения могли создать серьезные недостатки на протяжении миллионов лет.
| Wikispecies, где есть информация, относящаяся к Creodonta |
| Wikimedia Commons имеет медиа относится к Creodonta. |