
Войти
Гомосинаптическая пластичность - это один из видов синаптической пластичности. Гомосинаптическая пластичность зависит от входа, это означает, что изменения в силе синапсов происходят только на постсинаптических мишенях, специально стимулируемых пресинаптической мишенью. Следовательно, распространение сигнала от пресинаптической клетки локализовано.
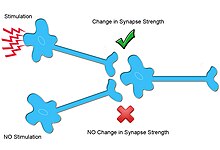 При гомосинаптической пластичности только нейроны, которые специфически иннервируются, претерпевают изменения в синаптической пластичности.
При гомосинаптической пластичности только нейроны, которые специфически иннервируются, претерпевают изменения в синаптической пластичности. Другой тип синаптической пластичности, гетеросинаптическая пластичность, не зависит от входа и отличается от гомосинаптической пластичности по многим механизмам.
Помимо специфичности входа, усиление синапса за счет гомосинаптической пластичности является ассоциативным, потому что оно тесно зависит от срабатывания пресинаптического и постсинаптического нейрона во времени. Эта ассоциативность увеличивает шансы срабатывания постсинаптического нейрона. Теоретически эти механизмы лежат в основе обучения и кратковременной памяти.
Дональд Хебб предположил, что усиление синаптических связей происходит из-за скоординированной активности между пресинаптическим концом и постсинаптическим дендритом. Согласно Хеббу, эти две клетки усилены, потому что их передача сигналов происходит вместе в пространстве и / или времени, что также известно как совпадающая активность. Этот постулат часто резюмируется как клетки, которые срабатывают вместе, соединяются вместе, что означает, что синапсы, у которых есть нейроны с совпадающими активациями, укрепляются, в то время как другие синапсы на этих нейронах остаются неизменными. Постулат Хебба предоставил концептуальную основу того, как синаптическая пластичность лежит в основе длительного хранения информации.
Изменения пластичности часто происходят через вставку или интернализацию рецепторов AMPA (AMPAR) в постсинаптическую мембрану синапса, претерпевающую изменение соединительной силы. Ca 2+ - это один сигнальный ион, который вызывает это изменение плотности рецептора AMPA, вызывая каскад биологических изменений внутри клетки. Чтобы вызвать долгосрочное усиление (LTP), Ca 2+ активирует CAMKII и PKC, вызывая фосфорилирование и вставку AMPAR, в то время как долговременная депрессия (LTD) происходит за счет активации Ca 2+ протеинфосфатаз, которые дефосфорилируют и вызывают интернализацию AMPAR.
Чтобы создать специфичные для входа изменения в силе синапсов, сигнал Ca 2+ должен ограничиваться определенными дендритными шипами. Дендритное ограничение Ca 2+ опосредуется несколькими механизмами. Внеклеточный Ca 2+ может проникать в позвоночник через рецепторы NMDA (NMDAR) и потенциал-управляемые каналы Ca 2+ (VGCC). И NMDAR, и VGCC сконцентрированы на дендритных шипах, опосредуя специфический для шипов приток Ca 2+. Внутриклеточные запасы Ca 2+ в эндоплазматическом ретикулуме и митохондриях также могут вносить вклад в ограниченную передачу сигналов в позвоночнике, хотя некоторые исследования не смогли найти доказательств этого. Клиренс Ca 2+ контролируется буферными белками, которые связываются с Ca 2+ и не дают ему просачиваться в другие шипы. Ограниченная диффузия Ca 2+ через шейку дендритного отростка также помогает изолировать его от определенных дендритов.
Другой механизм долгосрочной потенциации, зависящей от входов, является временным. NMDAR требуют как деполяризации, чтобы снять блокировку магния, так и активации глутамата, чтобы открыть свои каналы и обеспечить приток Ca 2+. Таким образом, LTP локализуется в местах, где каналы NMDA открываются активными синаптическими входами, которые высвобождают глутамат и вызывают деполяризацию постсинаптической клетки и не влияют на близлежащие неактивные синапсы.
Чтобы стабилизировать LTP и продлить его действие, новые белки, поддерживающие это изменение, синтезируются в ответ на стимуляцию в потенцирующем синапсе. Возникающая проблема состоит в том, как доставить специфические, вновь синтезированные белки к нужным входным специфическим синапсам, в которых они нуждаются. Два решения этой проблемы включают синаптическую маркировку и локальный синтез белка.
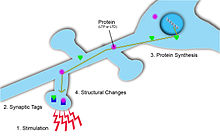 В нейроне синаптическая маркировка происходит в несколько этапов, чтобы предоставить информацию о синаптической пластичности.
В нейроне синаптическая маркировка происходит в несколько этапов, чтобы предоставить информацию о синаптической пластичности. Синаптические теги отмечают, где произошла синаптическая пластичность, и, таким образом, могут предоставить информацию о синаптической силе и потенциале долгосрочных пластических изменений. Метка является временной и включает большое количество белков, активируемых притоком Ca 2+ в постсинаптическую клетку. Кроме того, в зависимости от типа и величины синаптических изменений для маркировки используются разные белки. Например, когда пластические изменения приводят к длительной депрессии, применяется кальциневрин. И наоборот, когда пластичность приводит к длительному усилению, используется CaMKII. Для того, чтобы синаптическая пластичность была специфичной для входа, эти синаптические теги необходимы на постсинаптических мишенях, чтобы гарантировать локализацию синаптической потенциации. Эти метки позже инициируют синтез белка, который, в свою очередь, вызывает изменения синаптической пластичности в этих активированных нейронах.
Синтез белка в дендритах необходим для гомосинаптической пластичности. Деполяризация и связанная с этим активация рецепторов AMPA и NMDA в постсинаптической клетке вызывает эндоцитоз этих рецепторов. Для поддержания количества поверхностных рецепторов в синапсе необходим локальный синтез белка. Эти новые белки помогают стабилизировать структурные изменения, вызванные гомосинаптической пластичностью. Есть свидетельства наличия в дендритах рибосом, которые могут производить эти белки. Кроме того, есть доказательства наличия гранул РНК в дендритах, что указывает на присутствие вновь образованных белков. LTP может быть индуцирован из дендритов, отделенных от сомы постсинаптического нейрона-мишени. Напротив, LTP может быть заблокирован в этих дендритах блокаторами синтеза белка, такими как эндомиацин, что дополнительно указывает на место локального синтеза белка. Эти данные показывают, что для стабилизации и поддержания L-LTP необходим локальный синтез белка.