
Войти
Группа эквивалентности представляет собой набор неопределенных клеток, которые имеют одинаковый потенциал развития или способность принимать различные судьбы. Наше текущее понимание предполагает, что группы эквивалентности ограничиваются клетками одного происхождения, также известными как клетки-братья. Часто ячейки группы эквивалентности принимают разные судьбы друг от друга.
Группы эквивалентности предполагают разные потенциальные судьбы двумя общими, не исключающими друг друга способами. Один механизм, индукция, возникает, когда сигнал, исходящий извне группы эквивалентности, определяет подмножество наивных клеток. Другой режим, известный как латеральное ингибирование, возникает, когда сигнал в группе эквивалентности заставляет одну клетку принимать доминантную судьбу, в то время как другие в группе препятствуют этому. Во многих примерах групп эквивалентности как индукция, так и латеральное ингибирование используются для определения паттернов отдельных типов клеток.
Ячейки группы эквивалентности, которые не получают сигнал, принимают судьбу по умолчанию. С другой стороны, клетки, получившие сигнал, постигают разные судьбы. В определенный момент судьбы клеток в группе эквивалентности становятся необратимыми, поэтому они теряют свой мультипотентный потенциал. Ниже приведены примеры групп эквивалентности, изученных у нематод и асцидий.
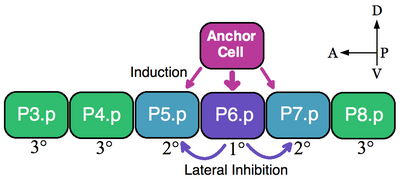 Модель для определения VPC (клетки P3.p-P8.p) у C. elegans. Индуктивный сигнал от якорной ячейки через LIN-3 определяет, что P6.p принимает 1 °. Затем боковой сигнал LIN-12 заставляет P5.p и P5.7 стать 2 °. Другие VPC не получают индуктивного сигнала и рассчитаны на 3 °. Передняя часть находится слева.
Модель для определения VPC (клетки P3.p-P8.p) у C. elegans. Индуктивный сигнал от якорной ячейки через LIN-3 определяет, что P6.p принимает 1 °. Затем боковой сигнал LIN-12 заставляет P5.p и P5.7 стать 2 °. Другие VPC не получают индуктивного сигнала и рассчитаны на 3 °. Передняя часть находится слева. Классическим примером группы эквивалентности являются клетки-предшественники вульвы (VPC) нематод. В Caenorhabditis elegans самооплодотворенные яйца выходят из тела через вульву. Этот орган развивается из подмножества клеток группы эквивалентности, состоящей из шести VPCs, P3.p-P8.p, которые лежат вентрально вдоль передне-задней оси. В этом примере одна вышележащая соматическая клетка, якорная клетка, побуждает соседние VPCs принимать судьбы вульвы на 1 ° (P6.p) и 2 ° (P5.p и P7.p). VPC, которые не индуцируются, образуют линию 3 ° (P3.p, P4.p и P8.p), которые образуют эпидермальные клетки, которые сливаются с большим синцитиальным эпидермисом (см. Изображение).
Шесть VPC образуются группа эквивалентности, потому что все шесть ячеек способны принять любую из доступных судьб (1 °, 2 ° и 3 °) в зависимости от их близости к якорной ячейке. Эксперименты по абляции показывают, что все VPC способны принимать судьбы вульвы. Напр., Если клетка P6.p, которая обычно становится 1 °, удаляется, тогда ближайший к якорной клетке VPC, P5.p или P7.p, принимает судьбу 1 °. Более того, если все VPCs разрушаются, кроме самой передней клетки P3.p, то якорная клетка определяет судьбу этой клетки 1 °. Однако, если якорная клетка погибает в отсутствие индуктивного сигнала, то все VPC принимают 3 ° клонирование по умолчанию.
Якорная клетка непосредственно индуцирует вульву судьбы путем секреции эпидермального фактора роста (EGF) -подобного лиганда LIN-3. Клетка P6.p получает сигнал LIN-3 через рецепторную тирозинкиназу LET-23 (P5.p и P7.p также получают LIN-3, но в меньшей степени). Активация LET-23 в P6.p приводит к активации LIN-12 (Notch ) в P5.p и P7.p. Экспериментальные данные показывают, что LIN-12 необходим и достаточен для формирования судьбы 2 °. Посредством латерального ингибирования LIN-12 предотвращает принятие клетками P5.p и P7.p линии 1 °. Таким образом, в этом примере и индуктивная передача сигналов EGF, и латеральные паттерны активации Notch являются группой эквивалентности VPC.
 Две двусторонние клетки-предшественники пигмента образуют группу эквивалентности, которая в конечном итоге взаимодействует, чтобы запечатать взаимоисключающие судьбы (отолит и глазок). Показан график развития эмбриона Halocythia roretzi (передняя часть слева). Клетки-предшественники пигмента (серые кружки) остаются эквивалентными до тех пор, пока не выравниваются после нейруляции. Через 21 час после оплодотворения определяют глазок (белый кружок) и отолит (черный кружок).
Две двусторонние клетки-предшественники пигмента образуют группу эквивалентности, которая в конечном итоге взаимодействует, чтобы запечатать взаимоисключающие судьбы (отолит и глазок). Показан график развития эмбриона Halocythia roretzi (передняя часть слева). Клетки-предшественники пигмента (серые кружки) остаются эквивалентными до тех пор, пока не выравниваются после нейруляции. Через 21 час после оплодотворения определяют глазок (белый кружок) и отолит (черный кружок). Личинки асцидий (морских брызг) содержат пару сенсорных пигментных клеток известный как отолит и глазок. Отолит используется для определения силы тяжести, а глазок реагирует на свет. Во время эмбриогенеза отолит и глазок развиваются из двух билатеральных эквивалентных предшественников. Как левая, так и правая клетка-предшественник пигмента имеет равную вероятность развития отолита или глазка. Решение принять любую судьбу определяется после закрытия нервной трубки на ранней стадии хвостовой почки (см. Изображение) посредством плохо определенного механизма индукции.
Во время нормального развития, после закрытия нервной трубки предшественники пигмента выстраиваются дорсально вдоль передне-задней оси нервной трубки. Какая бы клетка ни выровнялась вперед, она станет отолитом, а задняя клетка - глазком. В отсутствие межклеточных взаимодействий обе клетки развиваются в глазки, что является судьбой по умолчанию.
Чтобы выяснить, были ли судьбы отолита и глазка определяются в раннем эмбрионе или после того, как предшественники выравниваются во время закрытия нервной трубки, методы абляции и лекарственной обработки были использованы у видов асцидий Halocythia roretzi.
Клетки, меченные флуоресцеинизотиоцианат-декстраном (FDX), могут быть выборочно фотоаблируется флуоресцентным возбуждением. Когда одни клетки-предшественники пигмента, меченные FDX, подвергаются фотоабляции на стадии середины нейрулы (15 часов), другая почти всегда будет развиваться в глазок. Однако, если абляция проводится на поздней стадии хвостовой почки (22,5 часа), тогда оставшаяся клетка имеет равную вероятность стать отолитом или глазком.
Ингибирование деления клеток и морфогенеза с цитохалазин B представляет собой другой метод, используемый для определения того, когда указана группа эквивалентности предшественника пигмента. Обработка цитохалазином эмбрионов на ранней стадии хвостовой почки (17 часов), когда две двусторонние клетки все еще разделены, приводит к тому, что обе клетки становятся глазками. Когда лекарство применяли после того, как две клетки выровнялись по дорсальной средней линии, передняя клетка без исключения превратилась в отолит, а задняя клетка стала глазком. Оба эксперимента предполагают, что судьбы клеток-предшественников пигмента необратимо определяются примерно на стадии середины хвостовой почки (21 час).
Группы эквивалентности также были описаны в материнских клетках ганглия у кузнечика и O / P у пиявки. Как и другие экземпляры групп эквивалентности, клетки-потомки рождаются эквивалентными и становятся специфицированными посредством клеточных взаимодействий. Группы эквивалентности являются общей темой в развитии многих организмов из разных типов.