
Войти
| HLA-DQ8 (MHC Class II, DQ антиген клеточной поверхности) | |||
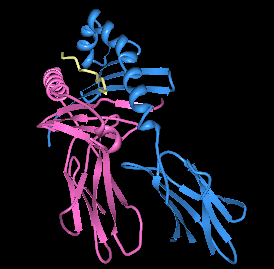 | |||
| Визуализация HLA -DQ8 с преобладающим инсулиновым пептидом в связывающем кармане. | |||
| Цис-гаплотип | Гаплотип | ||
| изоформа, | подтип | DQA1 | DQB1 |
| DQ αβ | DQ8.1 | * 0301 | * 0302 |
| DQ αβ | DQ8.1v | * 0302 | * 0302 |
| редкие гаплотипы | |||
| DQ αβ | DQ8.4 | * 0401 | * 0302 |
| DQ αβ | DQ8.5 | * 0503 | * 0302 |
. HLA-DQ8 (DQ8 ) - это лейкоцитарный антиген человека серотип в группе серотипа HLA-DQ (DQ). DQ8 представляет собой расщепленный антиген широкого антигена DQ3 широкого антигена. DQ8 определяется по распознаванию антителом β, и это обычно определяет продукт гена DQB1 * 0302.
DQ8 обычно связан с аутоиммунным заболеванием в человеческой популяции. DQ8 является второй наиболее распространенной изоформой, связанной с глютеновой болезнью, и DQ, наиболее связанной с диабетом 1 типа. DQ8 увеличивает риск ревматоидного артрита и связан с локусом первичного риска для РА, HLA-DR4. DR4 также играет важную роль при диабете 1 типа. В то время как гаплотип DQ8.1 связан с заболеванием, нет известной связи с гаплотипами DQB1 * 0305, DQ8.4 или DQ8.5 (см. Инфобокс) с аутоиммунным заболеванием; тем не менее, это может быть результатом недостаточного изучения популяций, которые их переносят, и очень низкой частоты.
DQ8.1 также отличается от других HLA частотами популяции. Обычно для антигенов MHC Class II у людей частота гаплотипов не превышает 40%. Например, в США самая высокая частота гаплотипа, гаплотип, кодирующий DQ6.2, составляет около 15%, что соответствует частотам фенотипа менее 30%. Частоты атипичных гаплотипов превышают 40%.
Для DQ8 самые высокие частоты гаплотипов достигают 80% в некоторых частях Центральной и Южной Америки, а частоты фенотипов приближаются к 90%. Это самая высокая частота фенотипа, наблюдаемая для любого фенотипа DR или DQ в человеческой популяции с большим отрывом.
| DQB1 * | DQ8 | DQ3 | DQ7 | Образец |
| аллеля | % | % | % | размер (N) |
| * 0302 | 66 | 23 | 4 | 6687 |
| * 0304 | 8 | 35 | 40 | 111 |
| * 0305 | 34 | 30 | 70 | |
| Красный указывает уровень «ложной» реакции у серотипов, не относящихся к DQ8 | ||||
| Связь аллелей с IMGT / HLA Databease в EBI | ||||
| freq | ||
| ref. | Население | (%) |
| Гватемала Майя | 48,1 | |
| Мексика Северный Леон Местисос | 22,5 | |
| США Южный Техас, латиноамериканцы | 20,6 | |
| Швеция | 18,7 | |
| Россия Сибирь Негидаль | 18,6 | |
| Россия Мурм анск Саоми | 18,5 | |
| Иордания Амман | 17,8 | |
| Самоа | 17,2 | |
| Англия европеоид | 16,4 | |
| Финляндия | 15,7 | |
| Франция | 14,5 | |
| Центральная Япония | 10,8 | |
| Греция Крит | 9,2 | |
| Испания Баскская долина Арратия | 6,7 | |
| Алжир Оран | 6,6 | |
| Китай Пекин и Сиань | 6,1 | |
| Эфиопия Амхара | 5,6 | |
| США ЮВ Афроамериканец | 4.9 | |
| США, Аляска, коренные жители юпиков | 3.8 | |
| Индия, северные индусы | 3.0 | |
| Зимбабве, Хараре Шона | 2.2 | |
| Россия Эскимосы Сибири | 0,9 | |
| Руанда Кигали Хуту и тутси | 0,5 | |
| PNG Восточное нагорье Горока | 0,0 | |
| Тунис Джерба Бербер | 0,0 |
Хотя ложная реакция с DQB1 * 0302 низкая, эффективность положительной реакции невысока, и существует риск ложного обнаружения DQB1 * 0305, что может привести к несовместимости. Для диагностики и подтверждения заболевания не существует известной связи DQB1 * 0305 ни с целиакией, ни с аутоиммунным диабетом. Таким образом, для DQ8 разумно использовать набор текста DQB1 с высоким разрешением.
| DQ | DQ | DQ | Freq | |||||
|---|---|---|---|---|---|---|---|---|
| Серотип | цис-изоформа | Подтип | A1 | B1 | % | |||
| DQ8 | α-β | 8,1 | 0301 | 0302 | 9. | 62 | ||
| 0302 | 0302 | 0. | 93 | |||||
| DQA1 * 0302 * 0303 не разрешен | ||||||||
DQ8 определяется распознаванием антителом β и осложняется тем фактом, что DQ8 хорошо распознает некоторые изоформы, кодируемые HLA-DQB1 * 03, частично частично или совсем плохо (см. серология) DQ β и β лучше всего распознаются как DQ8. Эти расщепленные антигены являются продуктами аллелей DQB1 * 0302 и DQB1 * 0305 соответственно.
| частота | ||
| ref. | Население | (%) |
| Ливан Кафар Зубиан | 5,9 | |
| Ливан Ниха эль-Шуфф | 2,5 | |
| Лакота Сиу | 2,1 | |
| Тунис | 2,0 | |
| Ливан Юхмур | 1,8 | |
| Гватемала Майя | 1,5 | |
| Марокко | 1,0 | |
| Пакистан | 1,0 | |
| Саудовская Аравия Гураят и Хайль | 0,8 | |
| Китай Лицзян Наси | 0,7 | |
| Россия Чувашия | 0,6 | |
| Тунис Матмата Бербер | 0,6 | |
| Чехия | 0,5 | |
| Индия Дели | 0,5 |
DQB1 * 0302 и чаще всего встречается в гаплотипе DQA1 * 0301: DQB1 * 0302, примерно в 10% случаев он встречается в гаплотипе DQA1 * 0302: DQB1 * 0302. DQB1 * 0302 почти всегда связаны с DR4, DRB1 * 0401, * 0402 и * 0404 у кавказцев. Первый и третий DRB1 наиболее сильно связаны с ревматоидным артритом.
Продукт гена DQB1 * 0305 немного более интенсивно реагирует с DQ8, чем DQ7, что обычно редко встречается в Европе и Северной Америке, за исключением нескольких коренных популяций. Уровни DQB1 * 0305, вероятно, выше, поскольку более ранние тесты не различали хорошо разные * 03.
β-цепи DQ8 объединяются с α-цепями, кодируемыми генетически связанными аллелями HLA-DQA1, с образованием гаплотипа цис - изоформы. Существует только одна общая цис-изоформа DQ8, потому что связанные аллели DQA1 * 03 (2) встречаются в большей части популяции, DQ8.1 является подавляющим большинством цис-изоформ DQ8. Редкий гаплотип DQA1 * 0503: DQB1 * 0302 обнаруживается ниже 1% всех гаплотипов DQ8 в Азии и Мезоамерике. Другой более редкий гаплотип, DQA1 * 0401: DQB1 * 0302
| Ссылка | DQA1 | DQB1 | Расчетная | |
|---|---|---|---|---|
| Население | *03 | * 0302 | DQ8.1 | |
| Лакандон Майан. (Мексика) | 79.0 | 77.9 | 77.9 | |
| Периджа-Юкпа. (Венесуэла) | 74,0 | 75,0 | 74,9 | |
| Майя. (Гватемала) | 48,1 | 47,6 | ||
| масатеканцы. (Мексика) | 48,5 | 48,5 | 47,5 | |
| Ламас. (Перу) | 45,2 | 44,7 | ||
| Дакота Сиу. (S. Дакота) | 52,1 | 45,0 | 44,5 | |
| Mixtec. (Оахака, Мексика) | 40,0 | 35,9 | 35,4 | |
| Лакота Сиу. (Южная Дакота) | 25,7 | 25,5 | ||
| Терена. (Бразилия) | 17,5 | 17,0 | ||
| Кавказ, Сан-Антонио. (США) | 11,7 | 11,7 | ||
| Кавказский. (США) | 18,5 | 10,5 | 10,5 | |
| афроамериканец. (SE. США) | 4,9 | 4,5 | ||
| Тлинглет. (Аляска, США) | 14,0 | 8,5 | 8,5 | |
| Эскимос. (Аляска, США) | 3,8 | 3,8 | ||
| Канончито Навахо. (Нью-Мексико, США) | 6,3 | 3,5 | 3,5 | |
| эскимосский. (Э. Гренландия) | 0,0 | 0,0 | 0,0 |
DQA1 * 0301: DQB1 * 0302 (DQ8.1) является наиболее распространенным подтипом DQ8, представляющим более 98% Несущая популяция DQ8. Нечасто DQA1 * 0302: DQB1 * 0302, но эта замена альфа-цепи, DQA1 ** 0301 по сравнению с * 0302, находится за пределами связывающей щели и, по-видимому, не изменяет функцию DQ8. DQ8.1 встречается почти повсеместно в каждой человеческой региональной популяции, но из-за своего уникального распространения становится объектом молекулярной антропологии. Есть 3 места, где частота гаплотипов повышена: Центральная и Южная Америка, Северо-Тихоокеанский регион и Северная Европа.
Глобальный узел для DQ8 находится в Центральной Америке и на севере Южной Америки, где он достигает самой высокой частоты для любого отдельного серотипа DQ, близкой к 90% частоте фенотипа (77 % частоты гаплотипа), и относительно высокая частота встречается у коренного населения Северной Америки и прибрежных районов Мексиканского залива и долины Миссисипи. Высокая частота DQ8 в северо-восточных регионах Южной Америки и низкая частота у коренных американцев более недавнего азиатского или сибирского происхождения позволяют предположить, что DQ8 был очень часто у самых ранних америндов. Характер распространения согласуется с недавними результатами мтДНК, предполагающими, что первые мигранты в Новый Свет поселились в низинных прибрежных районах, речных долинах и медленно двинулись вглубь страны, а последующие поселенцы переместились в высокогорные районы. DQ8 и DQ2.5 имеют много аналогичных функциональных сходств, и это смещение первого отстойника может быть причиной схожести. Исследования вируса Эпштейна-Барра и других белков предполагают, что оба белка являются кислыми (то есть пептидами с повышенным отрицательным зарядом) презентаторами пептидов (см. DQ8 для иллюстрации процесса презентации) и, возможно, были приспособлены для определенных стилей жизни охоты и собирательства, возможно, прибрежных собирателей.
| Ссылка | DQA1 | DQB1 | Расчетная | |
|---|---|---|---|---|
| Население | *03 | * 0302 | DQ8.1 | |
| Нивхи. (ННЕ. Сакалин И.) | 0,0 | 0 | ||
| Зорастра, Ядз. (Иран) | 20,8 | 0,8 | 0,8 | |
| Хотон. (Монголия) | 24,4 | 1,2 | 1,2 | |
| Маданг. (Папуа-Новая Гвинея) | 1,5 | 1,5 | 1,5 | |
| Яо. (Китай) | 2,6 | 2,6 | ||
| Наси. (Лицзян, Китай) | 2,7 | 2,7 | ||
| Айну. (Япония) | 3,0 | 3,0 | ||
| Шаньдун Хан. (Китай) | 3,1 | 3,1 | ||
| Халха. (Монголия) | 22,3 | 6,1 | 6,1 | |
| тайцы | 21,5 | 7,4 | 7,4 | |
| Кеть, Енисей. (Сибирь) | 14,7 | 8,4 | 8,4 | |
| Южная Корея | 10,3 | 10,3 | ||
| Японский | 10,8 | 10,8 | ||
| Нганасан. (Сибирь) | 39,6 | 11,4 | 11,4 | |
| Уйгары, Урумчи. (Китай) | 21,9 | 11,4 | 11,4 | |
| Казах | 11,4 | 11,4 | ||
| Негидаль. (Сибирь) | 50,0 | 18,6 | 18,6 | |
| Острова Кука. (Тихий океан) | 25,0 | 25,0 |
Хиатус из DQ8 в Северо-Восточная Сибирская Арктика, повышенные уровни в Амурской области и восточных турках
Уровни DQ8 на юго-западе к западному Тихоокеанскому региону находятся с переменной частотой гаплотипов, от 2 до 30%, и выравниваются примерно на 10% для Рюкюанов, японцев, корейцев, Амурская область и Северо-Западная часть Тихоокеанского региона упали до менее 1% в Нивхах. В настоящее время наблюдается перерыв DQ8 в регионе Аляска - Восточная Сибирь, и неясно, связано ли это с заменой, выбором или способом, которым прибыли первые американцы (то есть строго морским путем). Типы DR, связанные с DQ8: DRB1 * 0403, * 0404, * 0406, * 0407, * 0408, а * 0401 разделен между многими гаплотипами DQA1: B1. DQB1 * 0405 обычно ассоциируется с DQA1 * 0303: DQB1 * 04 и поэтому не включается в DRB1 * 0401 при оценке высокого разрешения. У DQ8 острова Кука был только один связанный гаплотип DR, предполагающий ограничивающее разнообразие интродукцию в регион, либо по маршруту TW- (Япония / Корея / Китай), либо через запад, например, у Bunun высокий DRB1 * 0403. Большая часть DRB1 * 04, по-видимому, была перераспределена из Восточной Азии из неизвестного источника, возможно, из Центральной Азии или Индии. Распределение можно сравнить с группами коренных жителей, такими как южноамериканцы. Три группы с высокими уровнями, когуи, сикуни и юкпа, имеют около 75% DQ8, доминирующим аллелем DRB1 * во 2 из 3 является * 0411 (Северный Китай = 0), но * 0407 (Рюкю, японец, манси). -Восточный Урал, китайцы-наси) и * 0403 (нганасаны, буряты, негидальцы, тунисцы, рюкю, Корея, айны). В Северной Америке DRB1 * 0404 и * 0407 более распространены, чем * 0403, а в Лакота Сиу B1 * 0411 встречается редко. Гаплотип DRB1 * 0404-DQ8 более распространен в Северо-Западной Азии и Северной Европе.
| Ссылка | DQA1 | DQB1 | Расчетное | |
|---|---|---|---|---|
| Население | *03 | * 0302 | DQ8.1 | |
| Ненец. (Северная Россия) | 40.9 | 20.9 | 20.0 | |
| Мурманск Саоми. (Россия) | 38.3 | 18,5 | 17,2 | |
| Архангельские Поморы. (Россия) | 22,6 | 17,1 | 16,4 | |
| Шведский | 24,2 | 18,7 | 16,1 | |
| европеоид. (Англия) | 23,7 | 16,4 | 15,9 | |
| Финляндия | 15,7 | 15,2 | ||
| CEPH Франции | 21,3 | 14,5 | 13,5 | |
| датчанин | 20,9 | 13,2 | 13,2 | |
| Голландский | 11,2 | 11,0 | ||
| Ирландский | 10,6 | |||
| Северо-западный славянский. (Россия) | 16,0 | 11,0 | 10,5 | |
| Немецкий | 10,5 | 9,9 | ||
| Русский | 8,9 | 7,2 | ||
| Кантабарский. (Испания) | 19,3 | 8,4 | 7,0 | |
| Испанский | 8,9 | 6,5 | ||
| Сардинский | 4,9 | 4,9 | ||
| Итальянский | 4,6 | 4,5 | ||
| Объединенные Арабские Эмираты | 0,83 | 0,8 | ||
| Иордания | 24,1 | 17,9 | 17,9 | |
| Африка | ||||
| ! Кунг | 36,7 | 36,7 | ||
| Хой | 14,9 | 13,9 | ||
| Оромо | 17,5 | 9,0 | 9,0 | |
| Марокко | 17,2 | 8,9 | 8,6 | |
| Тунис | 10,31 | 10,0 | 8,0 | |
| Амхара | 11,2 | 5,6 | 5,6 | |
| Пигмеи, известные как | 3,2 | 3,0 | ||
| Негроиды. (N. Африка) | 3,1 | 2,7 | ||
| Кэмерон | 11,5 | 1,5 | 1,2 | |
| Банту Конго | 0,6 | 0,5 | ||
| Габон | 7,5 | 0,5 | ||
| Буби | 17,5 | 0,0 | 0,0 | |
DQ8 также широко распространен в Северной Европе и часто встречается в немецко-скандинавско-уральской популяции к северу от Швейцарии. Гаплотипы HLA A-B предполагают, что миграция людей к востоку от Урала ответственна за DQ8, возможно, с востока, вплоть до Западной части Тихого океана. Высокий уровень DQ8 и DQ2.5 представляет большой интерес для заболеваний, опосредованных DQ, в Скандинавии и Северной Европе. DQ8 также встречается в Иберии, и в местах, где поток генов с востока на запад по другим генетическим маркерам не может быть подтвержден, и уровни в африканском или ближневосточном населении являются возможными источниками, Иберия имеет значительное уравновешивание A1 / B1, что предполагает независимые источники из Африки.
DQ8 вместе с несколькими другими гаплотипами, по-видимому, разделено на северо-запад / юго-восток в Евразии, а данные о DQ2.5 и других гаплотипах предполагают, что древнее население Центральной Азии было перемещены более поздней африканской миграцией. Есть много общих маркеров, обнаруженных во Франции, у немцев, датчан, шведов, тибетцев, у реки Амур, у японцев и корейцев, которые являются потенциальными индикаторами этого двустороннего распространения. Гаплотипы DQ8 обнаруживаются на высоких частотах в ! Kung, хотя можно ожидать большего количества DQ8 в Австронезии, он повсеместно распространен, если иногда низкие частоты, а иногда более высокие частоты (тайский язык). Путь распространения DQ8 в Новый Свет загадочен, несомненно, Япония и река Амур являются мощными источниками, но нельзя исключать наличие других перемещенных групп населения. Если способ передвижения был через коридор Берингии, как предлагает археолог, то очень низкая частота DQ8 в настоящее время является очень необычной находкой с точки зрения свидетельств полного перемещения в другие места в мире. Маркеры, общие для японских TW-аборигенов, имеют тенденцию к снижению частоты по мере приближения к Сибири, маркеры мтДНК уменьшаются в Курильской цепи. Во время периода Дзёмон в Японии предки Нинхвет / Айну должны были переместить и депрессию DQ8 по всей северной Японии, но упадок во всем регионе в некоторой степени необъясним, если не считать катастрофического климатического события между заселением Нового Света и Текущее время.
Альтернативная модель состоит в том, что было несколько источников DQ8 в населении северо-восточной Азии, некоторые источники были из Центральной Азии, а некоторые из индокитайского региона, некоторые из DQ8, обнаруженного на северо-западе Евразии, могли быть из примеси из источников Западно-Тихоокеанского региона и Центральной Азии, и были перемещены из более центральных регионов, но не из более восточных регионов.
Как и DQ2.5, DQ8 мог быть выбран для морских, прибрежных кормов и, в частности, для народов, адаптированных к климату / среде обитания на северной оконечности обитаемый западно-тихоокеанский рубеж на Последнем ледниковом максимуме. Выращивание Triticeae может привести к отрицательному отбору DQ8. Хотя существовало множество представителей видов Triticeae, подобных средневосточным диким Triticeae в Америке, и большое количество одомашненных растений в новом мире, ни один вид Triticeae, похоже, не был одомашнен в Новом Свете, и нет четких примеров в близкородственных племенах трав. Среди новых мировых видов трав в постколумбовские времена один вид Elymus был одомашнен для употребления в пищу человеком, а другой - в качестве пастырского сорта. Это можно интерпретировать двумя способами. Во-первых, этот уровень DQ8 отрицательно подавлял одомашнивание штаммов Triticeae. Во-вторых, отсутствие таких сортов, более подходящих, чем уже выведенные сорта, позволило DQ8 вырасти или остаться на высоком уровне, в то время как уровни DQ2.5 в NW при более длительной селекции упали, или немного того и другого. Большинство американских сортов было одомашнено к югу от Рио-Гранде (исключение составляют рис Каддо, кабачок техасский и т. Д.). Пшеница, особенно ячмень и рожь, являются предпочтительными сортами в более прохладном климате, в то время как Зеа более адаптивен к тропическому климату, а некоторые сорта относительно устойчивы к засухе, однако Зеа не хватает определенных аминокислот, которые необходимо дополнять другими продуктами, чтобы предотвратить недоедание. Близость неолитизации к экватору в Новом Свете может иметь прямое отношение к неочевидному негативному отбору DQ8 по сравнению с неолитизацией Западной Евразии.
 Отображение HLA-DQ8 с инсулиновым пептидом в связывающем кармане, смотрящим вниз на связывающую бороздку
Отображение HLA-DQ8 с инсулиновым пептидом в связывающем кармане, смотрящим вниз на связывающую бороздку В Европе DQ8 связан с типом 1 диабет и глютеновая болезнь (также известная как глютеновая болезнь). Самым высоким фактором риска диабета 1 типа является фенотип HLA DQ8 / DQ2.5. В некоторых частях Восточной Скандинавии как DQ2.5, так и DQ8 являются высокими повышениями частоты позднего начала диабета типа I и неоднозначного диабета типа I / II. DQ8 также обнаружен у многих коренных народов Азии, он был обнаружен на раннем этапе у бедуинского населения Аравии, где DQ2.5 часто отсутствует, и в этих случаях DQ8 является исключительно ассоциированным HLA при глютеновой болезни.
Однако в Соединенных Штатах, похоже, наблюдается сдвиг в риске аутоиммунных заболеваний для иммигрантов из Мексики. Повышенная иммунореактивность латиноамериканцев в Хьюстоне, по-видимому, связана с DR4-DQ8. Гаплотип может вызвать самый высокий риск ревматоидного артрита.
В Японии DQ3 (DQ7, DQ8, DQ9) связан с миастенией у женщин с ранним началом, хотя не кажется, что DQ8 играет большую роль, между миастенией есть сходство gravis в Японии и обнаруженный в латиноамериканской популяции Хьюстона, при этом DQ8 ассоциировался с более молодыми женщинами по сравнению с ассоциациями всех других типов HLA DQ. В Японии наблюдается рост глютеновой болезни, и очевидно, что причиной этого являются изменения в питании, но в Японии также нет DQ2,5, а уровни DQ8 умеренные.
Многие заболевания, связанные с DQ8, имеют двойную связь с DR4, а некоторые DR4 (* 0405) имеют независимую и зависимую ассоциацию риска с DQ8, например, с диабетом 1 типа.
Целиакия
Тип 1 Диабет