
Войти
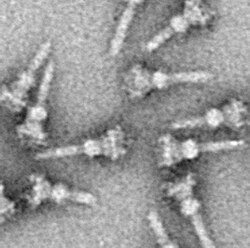 Изображение на просвечивающем электронном микроскопе изолированных комплексов игл T3SS из Salmonella typhimurium
Изображение на просвечивающем электронном микроскопе изолированных комплексов игл T3SS из Salmonella typhimurium Система секреции третьего типа (часто обозначаемая как система секреции типа III и сокращенно TTSS или T3SS, также называемая инъектисомой) - это белковый придаток, обнаруженный у нескольких грамотрицательных бактерий.
У патогенных бактерий игольчатая структура используется в качестве сенсорного зонда для обнаружения присутствия эукариотических организмов и выделения белков, которые помогают бактериям их инфицировать. Секретируемые эффекторные белки секретируются непосредственно из бактериальной клетки в эукариотическую (хозяйскую) клетку, где они оказывают ряд эффектов, которые помогают патогену выжить и избежать иммунного ответа.
Термин «система секреции типа III» был придуман в 1993 году. Эта система секреции отличается, по крайней мере, от пяти других систем секреции, обнаруженных у грамотрицательных бактерий. Многие бактерии, ассоциированные с животными и растениями, обладают похожими T3SS. Эти T3SS похожи в результате дивергентной эволюции, и филогенетический анализ поддерживает модель, в которой грамотрицательные бактерии могут переносить кассету гена T3SS горизонтально другим видам. Наиболее изученными T3SS являются виды Shigella (вызывает бактериальную дизентерию ), Salmonella ( брюшной тиф ), Escherichia coli ( кишечная флора, некоторые штаммы вызывают пищевое отравление ), Vibrio ( гастроэнтерит и диарея ), Burkholderia ( сап ), Yersinia ( чума )., Хламидиоз ( болезнь, передающаяся половым путем ), псевдомонаду (поражает людей, животных и растения ) и патогены растений Erwinia, Ralstonia и Xanthomonas, а также растительный симбионт Rhizobium.
T3SS состоит примерно из 30 различных белков, что делает его одной из самых сложных систем секреции. Его структура имеет много общего с бактериальными жгутиками (длинные жесткие внеклеточные структуры, используемые для подвижности ). Некоторые из белков, участвующих в T3SS, имеют гомологию аминокислотной последовательности с жгутиковыми белками. Некоторые из бактерий, обладающих T3SS, также имеют жгутики и подвижны (например, Salmonella), а некоторые нет (например, Shigella). С технической точки зрения, секреция типа III используется как для секреции белков, связанных с инфекцией, так и для жгутиковых компонентов. Однако термин «секреция типа III» используется в основном в отношении инфекционного аппарата. Жгутик бактерий имеет общего предка с системой секреции типа III.
T3SS необходимы для патогенности (способности инфицировать) многих патогенных бактерий. Дефекты T3SS могут сделать бактерию непатогенной. Было высказано предположение, что некоторые неинвазивные штаммы грамотрицательных бактерий потеряли T3SS, потому что энергетически дорогостоящая система больше не используется. Хотя традиционные антибиотики были эффективны против этих бактерий в прошлом, постоянно появляются устойчивые к антибиотикам штаммы. Понимание того, как работает T3SS, и разработка лекарств, специально нацеленных на него, стали важной целью многих исследовательских групп по всему миру с конца 1990-х годов.
| Система секреции типа III | |
|---|---|
 Игольчатый комплекс T3SS Игольчатый комплекс T3SS | |
| Идентификаторы | |
| Условное обозначение | T3SS |
| TCDB | 1.B.22 |
| OPM суперсемейство | 348 |
| Белок OPM | 5ткк |
Отличительной чертой T3SS является игла (в более общем смысле, комплекс иглы ( NC) или аппарат T3SS ( T3SA); также называемый инъектисомой, когда АТФаза исключена; см. Ниже). Бактериальные белки, которые необходимо секретировать, проходят из бактериальной цитоплазмы через иглу непосредственно в цитоплазму хозяина. Две цитоплазмы разделяют три мембраны : двойные мембраны (внутренняя и внешняя мембраны) грамотрицательных бактерий и эукариотическая мембрана. Игла обеспечивает плавный проход через эти высокоселективные и почти непроницаемые мембраны. У одной бактерии может быть несколько сотен игольчатых комплексов, распределенных по ее мембране. Было высказано предположение, что комплекс иглы является универсальным признаком всех T3SS патогенных бактерий.
Комплекс иглы начинается в цитоплазме бактерии, пересекает две мембраны и выступает из клетки. Часть, закрепленная в мембране, является основанием (или базальным телом) T3SS. Внеклеточная часть - игла. Так называемый внутренний стержень соединяет иглу с основанием. Сама игла, хотя и является самой большой и наиболее заметной частью T3SS, состоит из множества единиц одного белка. Таким образом, большинство различных белков T3SS - это те, которые образуют основу, и те, которые секретируются в хозяина. Как упоминалось выше, комплекс игл имеет сходство с бактериальными жгутиками. Более конкретно, основание комплекса игл структурно очень похоже на основание жгутика; сама игла аналогична крючку жгутика - структуре, соединяющей основание с нитью жгутика.
Основание состоит из нескольких круглых колец и является первой структурой, построенной в новом комплексе игл. Когда основа готова, она служит машиной для выделения внешних белков (иглы). После завершения всего комплекса система переключается на секретирование белков, которые предназначены для доставки в клетки-хозяева. Предполагается, что игла построена снизу вверх; единицы мономерного белка иглы накладываются друг на друга, так что единица на кончике иглы добавляется последней. Игла субъединица является одним из самых маленьких T3SS белков, измерение на уровне около 9 K Da. Каждая игла состоит из 100-150 субъединиц.
Игла T3SS имеет длину около 60-80 нм и внешнюю ширину 8 нм. Он должен иметь минимальную длину, чтобы другие внеклеточные бактериальные структуры (например, адгезины и липополисахаридный слой) не мешали секреции. Отверстие иглы имеет диаметр 3 нм. Большинство свернутых эффекторных белков слишком велики, чтобы пройти через отверстие иглы, поэтому большая часть секретируемых белков должна проходить через иглу в развернутом виде, и эту задачу выполняет АТФаза в основании структуры.
 Схема отдельных субструктур комплекса игл Salmonella typhimurium
Схема отдельных субструктур комплекса игл Salmonella typhimurium Белки T3SS можно разделить на три категории:
Большинство генов T3SS расположены в оперонах. Эти опероны расположены на бактериальной хромосоме у некоторых видов и на специальной плазмиде у других видов. Сальмонелла, например, имеет хромосомную область, в которой сосредоточено большинство генов T3SS, так называемый остров патогенности сальмонелл ( SPI). Shigella, с другой стороны, имеет большую плазмиду вирулентности, на которой расположены все гены T3SS. Важно отметить, что многие островки патогенности и плазмиды содержат элементы, которые делают возможным частый горизонтальный перенос генов острова / плазмиды новому виду.
Эффекторные белки, которые должны секретироваться через иглу, должны распознаваться системой, поскольку они плавают в цитоплазме вместе с тысячами других белков. Распознавание осуществляется с помощью сигнала секреции - короткой последовательности аминокислот, расположенной в начале ( N-конце ) белка (обычно в пределах первых 20 аминокислот), которую комплекс иглы способен распознать. В отличие от других систем секреции, сигнал секреции белков T3SS никогда не отщепляется от белка.
Контакт иглы с клеткой-хозяином запускает секрецию T3SS; Об этом спусковом механизме известно немного (см. ниже). Секреция также может быть вызвана снижением концентрации ионов кальция в среде для выращивания (для Yersinia и Pseudomonas ; выполняется путем добавления хелатора, такого как EDTA или EGTA ) и путем добавления ароматического красителя Конго красный в среду для выращивания (для Shigella). например. Эти и другие методы используются в лабораториях для искусственной индукции секреции III типа.
Индукция секреции с помощью внешних сигналов, отличных от контакта с клетками-хозяевами, также имеет место in vivo у инфицированных организмов. Бактерии воспринимают такие сигналы, как температура, pH, осмолярность и уровень кислорода, и используют их, чтобы «решить», активировать ли их T3SS. Например, сальмонелла может лучше размножаться и проникать в подвздошную кишку, а не в слепую кишку кишечника животных. Бактерии могут знать, где они находятся, благодаря различным ионам, присутствующим в этих областях; подвздошная кишка содержит формиат и ацетат, а слепая кишка - нет. Бактерии ощущают эти молекулы, определяют, что они находятся в подвздошной кишке, и активируют свой секреторный аппарат. Молекулы, присутствующие в слепой кишке, такие как пропионат и бутират, являются негативным сигналом для бактерий и подавляют секрецию. Холестерин, липид, содержащийся в мембранах большинства эукариотических клеток, способен вызывать секрецию у Shigella.
Перечисленные выше внешние сигналы регулируют секрецию либо напрямую, либо через генетический механизм. Известно несколько факторов транскрипции, регулирующих экспрессию генов T3SS. Некоторые шапероны, связывающие эффекторы T3SS, также действуют как факторы транскрипции. Был предложен механизм обратной связи: когда бактерия не секретирует, ее эффекторные белки связываются с шаперонами и плавают в цитоплазме. Когда начинается секреция, шапероны отделяются от эффекторов, и последние секретируются и покидают клетку. Затем одиночные шапероны действуют как факторы транскрипции, связываясь с генами, кодирующими их эффекторы, и индуцируют их транскрипцию и тем самым производство большего количества эффекторов.
Структуры, аналогичные инъекциомам Type3SS, были предложены для заклепки грамотрицательных бактериальных наружных и внутренних мембран, чтобы помочь высвободить везикулы внешней мембраны, нацеленные на доставку бактериального секрета к эукариотическим клеткам-хозяевам или другим клеткам-мишеням in vivo.
Эффекторы T3SS входят в комплекс иглы в основании и продвигаются внутрь иглы к клетке-хозяину. Точный путь проникновения эффекторов в организм хозяина в основном неизвестен. Ранее предполагалось, что игла сама по себе способна прокалывать отверстие в мембране клетки-хозяина; эта теория была опровергнута. Теперь ясно, что некоторые эффекторы, вместе называемые транслокаторами, секретируются первыми и продуцируют поры или канал ( транслокон) в мембране клетки-хозяина, через которые могут проникать другие эффекторы. Мутировавшие бактерии, у которых отсутствуют транслокаторы, способны секретировать белки, но не могут доставлять их в клетки-хозяева. Обычно каждый T3SS включает три транслокатора. Некоторые транслокаторы выполняют двойную роль; после того, как они участвуют в порообразовании, они проникают в клетку и действуют как истинные эффекторы.
Эффекторы T3SS манипулируют клетками-хозяевами несколькими способами. Самый поразительный эффект - это стимуляция поглощения бактерии клеткой-хозяином. Многие бактерии, обладающие T3SS, должны проникать в клетки-хозяева, чтобы реплицировать и распространять инфекцию. Эффекторы, которые они вводят в клетку-хозяин, побуждают хозяина поглотить бактерию и практически «съесть» ее. Для этого бактериальные эффекторы манипулируют аппаратом полимеризации актина клетки-хозяина. Актин является компонентом цитоскелета, а также участвует в подвижности и изменении формы клеток. Благодаря своим эффекторам T3SS бактерия может использовать собственный механизм клетки-хозяина для собственной выгоды. Как только бактерия попадает в клетку, она способна более легко секретировать другие эффекторы, может проникать в соседние клетки и быстро инфицировать всю ткань.
Было также показано, что эффекторы T3SS влияют на клеточный цикл хозяина, и некоторые из них способны вызывать апоптоз. Одним из наиболее изученных эффекторов T3SS является IpaB из Shigella flexneri. Он выполняет двойную роль как транслокатор, создавая поры в мембране клетки-хозяина, и как эффектор, оказывая множественные пагубные эффекты на клетку-хозяина. Было продемонстрировано, что IpaB вызывает апоптоз в макрофагах - клетках иммунной системы животных - после их поглощения ими. Позже было показано, что IpaB достигает этого за счет взаимодействия с каспазой 1, основным регуляторным белком в эукариотических клетках.
Другим хорошо охарактеризованным классом эффекторов T3SS являются эффекторы, подобные активаторам транскрипции ( эффекторы TAL ) из Xanthomonas. При введении в растения эти белки могут проникать в ядро растительной клетки, связывать промоторные последовательности растений и активировать транскрипцию генов растений, которые способствуют бактериальной инфекции. Недавно было продемонстрировано, что распознавание эффекторной ДНК TAL включает простой код, и это значительно улучшило понимание того, как эти белки могут изменять транскрипцию генов в клетках растения-хозяина.
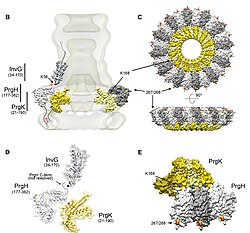 Топология и организация игольного комплекса сальмонелл.
Топология и организация игольного комплекса сальмонелл. Сотни статей о T3SS были опубликованы с середины девяностых. Однако многие вопросы, касающиеся системы, остаются нерешенными:
 Жгутик грамотрицательных бактерий. Кольца основы очень похожи на кольца со сложной иглой, хотя наличие С-образного кольца в комплексе с иглой не доказано. Жгутиковый крючок гомологичен игле T3SS.
Жгутик грамотрицательных бактерий. Кольца основы очень похожи на кольца со сложной иглой, хотя наличие С-образного кольца в комплексе с иглой не доказано. Жгутиковый крючок гомологичен игле T3SS. С начала 1990-х годов новые белки T3SS постоянно обнаруживаются у разных видов бактерий. Аббревиатуры были даны независимо для каждой серии белков в каждом организме, и названия обычно мало что говорят о функции белка. Позднее было показано, что некоторые белки, независимо открытые в разных бактериях, гомологичны ; исторические названия, однако, в основном были сохранены, что может вызвать путаницу. Например, белки SicA, IpgC и SycD являются гомологами Salmonella, Shigella и Yersinia, соответственно, но последняя буква («серийный номер») в их названии этого не показывает.
Ниже приводится краткое изложение наиболее распространенных названий серий белков у нескольких видов, содержащих T3SS. Обратите внимание, что эти названия включают белки, которые образуют механизм T3SS, а также секретируемые эффекторные белки :
После этих сокращений следует буква или цифра. Буквы обычно обозначают «порядковый номер», либо хронологический порядок открытия, либо физический порядок появления гена в опероне. Числа, более редкий случай, обозначают молекулярную массу белка в кДа. Примеры: IpaA, IpaB, IpaC; MxiH, MxiG, MxiM; Спа9, Спа47.
Во всех T3SS присутствуют несколько ключевых элементов: мономер иглы, внутренний стержень иглы, кольцевые белки, два транслокатора, белок кончика иглы, белок-линейка (который, как считается, определяет длину иглы; см. Выше) и АТФазы, которая поставляет энергию для секреции. В следующей таблице показаны некоторые из этих ключевых белков у четырех бактерий, содержащих T3SS:
| ↓ Функция / Род → | Шигелла | Сальмонелла | Иерсиния | Эшерихия |
|---|---|---|---|---|
| Игольчатый мономер | MxiH | PrgI | YscF | EscF |
| Внутренний стержень | MxiI | PrgJ | YscI | EscI |
| Белок кончика иглы | IpaD | SipD | LcrV | EspA |
| Транслокатор | IpaB | SipB | YopB | EspD |
| Транслокатор | IpaC | SipC | YopD | EspB |
| Шаперон для двух транслокаторов | IpgC | SicA | SycD | CesD |
| АТФаза | Спа47 | InvC | YscN | SepB (EscN) |
| Линейка белка | Спа32 | InvJ | YscP | Orf16 |
| Выключатель | Спа40 | СПАС | YscU | EscU |
| Привратник | MxiC | InvE | YopN (TyeA) | SepL |
Выделение крупных, хрупких, гидрофобных мембранных структур из клеток было проблемой в течение многих лет. Однако к концу 1990-х годов было разработано несколько подходов к изоляции T3SS NC. В 1998 г. были выделены первые НК из Salmonella typhimurium.
Для изоляции бактерии выращивают в большом объеме жидкой питательной среды до достижения логарифмической фазы. Затем их центрифугируют ; надосадочную жидкость (среда) отбрасывают, а осадок (бактерии) ресуспендировали в буфере для лизиса, как правило, содержащий лизоцим, а иногда и моющее средство, такие как LDAO или Тритон Х-100. Этот буфер разрушает клеточную стенку. После нескольких циклов лизиса и промывки открытые бактерии подвергают серии ультрацентрифугирования. Эта обработка обогащает крупные макромолекулярные структуры и удаляет более мелкие клеточные компоненты. Необязательно, конечный лизат подвергают дальнейшей очистке градиентом плотности CsCl.
Дополнительный подход к дальнейшей очистке использует аффинную хроматографию. Рекомбинантные белки T3SS, несущие белковые метки (например, гистидиновые метки ), производятся путем молекулярного клонирования, а затем вводятся ( трансформируются ) в исследуемые бактерии. После первоначального выделения NC, как описано выше, лизат пропускают через колонку, покрытую частицами с высоким сродством к метке (в случае гистидиновых меток: ионы никеля ). Меченый белок остается в колонке, а вместе с ним и весь комплекс иглы. С помощью таких методов можно достичь высокой степени чистоты. Эта чистота важна для многих деликатных анализов, которые использовались для определения характеристик NC.
Эффекторы типа III были известны с начала 1990-х годов, но способ их доставки в клетки-хозяева оставался полной загадкой. Гомология между многими жгутиковыми белками и белками T3SS заставила исследователей подозревать существование внешней структуры T3SS, подобной жгутикам. Идентификация и последующее выделение структуры иглы позволили исследователям:
Как и почти все белки, визуализация НК T3SS возможна только с помощью электронной микроскопии. Первые изображения NC (1998) показали игольчатые структуры, выступающие из клеточной стенки живых бактерий и плоские, двумерные изолированные NC. В 2001 г. изображения NC из Shigella flexneri были проанализированы в цифровом виде и усреднены для получения первой полу-3D-структуры NC. Спиральная структура NC из Shigella flexneri была определена с разрешением 16 Å с помощью дифракции рентгеновских лучей в 2003 году, а год спустя была опубликована трехмерная структура 17 Å NC из Salmonella typhimurium. Последние достижения и подходы позволили получить трехмерные изображения NC с высоким разрешением, дополнительно прояснив сложную структуру NC.
Многочисленные белки T3SS были кристаллизованы на протяжении многих лет. К ним относятся структурные белки NC, эффекторы и шапероны. Первой структурой мономера игольчатого комплекса была структура ЯМР BsaL из "Burkholderia pseudomallei", а затем кристаллическая структура MixH из Shigella flexneri, которые были разрешены в 2006 году.
В 2012 году сочетание производства рекомбинантных игл дикого типа, твердотельного ЯМР, электронной микроскопии и моделирования Rosetta выявило супрамолекулярные границы раздела и, в конечном итоге, полную атомную структуру иглы Salmonella typhimurium T3SS. Было показано, что 80-остатки PrgI субъединицы образуют правую винтовую сборку с примерно 11 субъединиц на два оборота, подобно тому, что часть жгутика из Salmonella Typhimurium. Модель также выявила протяженный аминоконцевой домен, который расположен на поверхности иглы, в то время как высококонсервативный карбоксильный конец указывает на просвет.
Несколько методов были использованы для идентификации набора белков, составляющих T3SS. Изолированные комплексы игл могут быть разделены с помощью SDS-PAGE. Полосы, которые появляются после окрашивания, можно индивидуально вырезать из геля и анализировать с помощью секвенирования белков и масс-спектрометрии. Структурные компоненты NC могут быть отделены друг от друга (например, игольчатая часть от основной части), и путем анализа этих фракций можно определить белки, участвующие в каждой из них. В качестве альтернативы, изолированные NC могут быть непосредственно проанализированы масс-спектрометрией без предварительного электрофореза, чтобы получить полную картину протеома NC.
Исследователи манипулировали T3SS у многих бактерий. Наблюдение за влиянием отдельных манипуляций может быть использовано для понимания роли каждого компонента системы. Примеры манипуляций:
Манипуляции с компонентами T3SS могут влиять на несколько аспектов бактериальной функции и патогенности. Примеры возможных влияний:
Было обнаружено несколько соединений, которые ингибируют T3SS у грамотрицательных бактерий, включая гуадиномины, которые естественным образом вырабатываются видами Streptomyces. Были разработаны моноклональные антитела, которые также ингибируют T3SS. Было показано, что Aurodox, антибиотик, способный ингибировать трансляцию белков T3SS, способен предотвращать эффекторы T3SS in vitro и на животных моделях.