
Войти
 Палеонтологическое древо позвоночных Геккеля (ок. 1879). Эволюционная история вида описывается как «дерево » с множеством ветвей, возникающих из одного ствола. Хотя дерево Геккеля несколько устарело, оно ясно иллюстрирует принципы, которые могут затенить более сложные современные реконструкции.
Палеонтологическое древо позвоночных Геккеля (ок. 1879). Эволюционная история вида описывается как «дерево » с множеством ветвей, возникающих из одного ствола. Хотя дерево Геккеля несколько устарело, оно ясно иллюстрирует принципы, которые могут затенить более сложные современные реконструкции. Хронология эволюции человека очерчивает основные события в эволюционной линии современного человеческого вида., Homo sapiens на протяжении истории жизни, начиная примерно 4,2 миллиарда лет назад и заканчивая недавней эволюцией внутри H. sapiens во время и после Последний ледниковый период.
Он включает краткие объяснения различных таксономических рангов в человеческой родословной. Хронология отражает основные взгляды в современной таксономии, основанные на принципе филогенетической номенклатуры ; в случае открытых вопросов без четкого консенсуса кратко описываются основные конкурирующие возможности.
Табличный обзор таксономический рейтинг для Homo sapiens (с оценками возраста для каждого ранга) показан ниже.
| Ранг | Имя | Обычное имя | Миллионы лет назад (начало) | |
|---|---|---|---|---|
| Жизнь | 4200 | |||
| Архея | ||||
| Домен | Eukaryota | Eukaryotes (слизевики и родственные им) | 2100 | |
| Podiata | ||||
| Unikonts | ||||
| Obazoa | ||||
| Opisthokonts | Holozoa + Fungi сл | 1,300 | ||
| Holozoa | 1,100 | |||
| Filozoa | Choanozoa + Filasterea | |||
| Choanozoa | Choanoflagelates + Животные | 900 | ||
| Королевство | Animalia | Животные | 610 | |
| Subkingdom | Eumetazoa | |||
| Parahoxozoa | ||||
| Bilateria | Триплобласты / черви | 560 | ||
| Nephrozoa | ||||
| Deuterstomes | ||||
| Тип | Chordata | Chordata (позвоночные и близкородственные беспозвоночные) | 530 | |
| Olfactores | ||||
| Subphylum | Vertebrata | Рыба / позвоночные | 505 | |
| Infraphylum | Gnathostomata | Рыба с челюстями | 460 | |
| Teleostomi | Костяная рыба | 420 | ||
| Sarcopterygii | Рыбные плавники с лепестками | |||
| Суперкласс | Tetrapoda | Tetrapods (животные с четырьмя конечностями) | 395 | |
| Amniota | Amniotes (полностью наземные четвероногие, чьи яйца «снабжены амниосом» ) | 340 | ||
| Synapsida | Прото-млекопитающие | 308 | ||
| Therapsid | Конечности под телом и другие признаки млекопитающих | 280 | ||
| Класс | Mammalia | Млекопитающие | 220 | |
| Подкласс | Theria | Млекопитающие, которые рождают живых детенышей (т. Е. Не откладывают яйца) | 160 | |
| Infraclass | Eutheria | Placental млекопитающие (т. е. не сумчатые) | 125 | |
| Magnorder | Boreoeutheria | Supraprimates, (большинство) копытных млекопитающих, (большинство) плотоядных млекопитающих, киты и летучие мыши | 124–101 | |
| Надотряд | Euarchontoglires | Супраприматы: приматы, колуги, землеройки, грызуны и кролики | 100 | |
| Грандордер | Euarchonta | Приматы, колуги и землеройки | 99–80 | |
| Зеркалоотряд | Приматоморфа | Приматы и колуго | 79.6 | |
| Отряд | Приматы | Приматы / Plesiadapiformes | 75 | |
| Подотряд | Haplorrhini | «Сухоносые» (буквально «простоносые») приматы: долгопяты и обезьяны (вкл. обезьяны ) | 63 | |
| Инфраотряд | Simiiformes | обезьяны (включая обезьян) | 40 | |
| Parvorder | Catarrhini | «низконосые» приматы: обезьяны и обезьяны старого мира | 30 | |
| Надсемейство | Hominoidea | Обезьяны: большие обезьяны и малые обезьяны (гиббоны) | 28 | |
| Семейство | Hominidae | человекообразные обезьяны : люди, шимпанзе, гориллы и орангутаны - гоминиды | 20–15 | |
| Подсемейство | Homininae | Люди, шимпанзе и гориллы (африканские обезьяны) | 14–12 | |
| Племя | Гоминини | Включает как человека, так и пан (шимпанзе), но не гориллу. | 10–8 | |
| Подтриба | Hominina | Род Homo и близкие человеческие родственники и предки после отделения от Пана - гоминины | 8–4 | |
| (Род) | Ardipithecus сл | 6-4 | ||
| (Род) | Australopithecus | 3 | ||
| Род | Homo (H. Habilis) | Люди | 2,5 | |
| ( Виды) | H. Erectus s.l. | |||
| (Вид) | Х. heidelbergensis s.l. | |||
| Виды | Homo sapiens | Анатомически современные люди | 0,8–0,3 |
| Дата | Событие |
|---|---|
| 4,1 Ga. (миллиард. лет назад) | Появляется самая ранняя жизнь. |
| 3.9 Ga | Появляются клетки, похожие на прокариот. |
| 3,5 Ga | Это знаменует первое появление кислородного фотосинтеза и, следовательно, первое появление больших количеств атмосферного кислорода на Земле. |
| 2,5 млрд лет | Первые организмы, использующие кислород. К 2400 млн. Лет назад, во время так называемого Великого события оксигенации, предкислородные анаэробные формы жизни были уничтожены продуцентами кислорода. |
| 2.1 Ga | Появляются более сложные клетки: эукариоты. |
| 1.2 Ga | Половое размножение развивается, что приводит к более быстрой эволюции, при которой гены смешиваются в каждом поколении, что обеспечивает большее разнообразие для последующего выбора. |
| 0,9 Ga |  Choanoflagellate Choanoflagellate choanoflagellates могут быть похожи на предков всего животного царства, и, в частности, они могут быть прямыми предками губок. Proterospongia (представители Choanoflagellata) являются лучшими живыми примерами того, как мог выглядеть предок всех животных. Они живут в колониях и демонстрируют примитивный уровень клеточной специализации для различных задач. |
| Дата | Событие |
|---|---|
| 700–660 Ma |  Dickinsonia costata из эдиакарской биоты, 635–542 млн лет назад, возможно раннее член Animalia. Dickinsonia costata из эдиакарской биоты, 635–542 млн лет назад, возможно раннее член Animalia. Urmetazoan : Первые окаменелости, которые могут представлять животных, появляются в скалах возрастом 665 миллионов лет в формации Trezona из Южная Австралия. Эти окаменелости интерпретируются как ранние губки. Отделение от линии Porifera (sponges ). Eumetazoa / Diploblast : отделение от линии Ctenophora («гребешки»). Planulozoa / ParaHoxozoa : отделение от линий Placozoa и Cnidaria. Почти все книдарии обладают нервами и мышцами. Поскольку они являются простейшими животными, чтобы ими обладать, их прямые предки, вероятно, были первыми животными, которые использовали вместе нервы и мышцы. Книдарианцы также являются первыми животными с реальным телом определенной формы и формы. Они имеют радиальную симметрию. Первые глаза появились в это время. |
| 570–550 млн лет назад |  Proporus sp., A ксенакоеломорф. Proporus sp., A ксенакоеломорф.Urbilaterian : Bilateria / Triploblasts, Nephrozoa (555 млн лет назад), последний общий предок протостомов (включая линии членистоногих [насекомых, ракообразных] и платизоанов [плоских червей]) и deuterostomes (включая линию позвоночных [человеческих]). Самое раннее развитие мозга и двусторонней симметрии. Архаичными представителями этой стадии являются плоские черви, простейшие животные, органы которых формируются из трех зародышевых листков. |
| 541 млн лет |  A морского огурца (Actinopyga echinites ), демонстрируя питающие щупальца и трубчатые ножки. A морского огурца (Actinopyga echinites ), демонстрируя питающие щупальца и трубчатые ножки.Наиболее известные типы животных появились в летописи окаменелостей как морские виды во время кембрийского взрыва. Deuterostomes, последнего общего предка хордовые [человеческие] линии, иглокожие (морские звезды, морские ежи, морские огурцы и т. д.) и Hemichordata (желудевые черви и граптолиты ). архаичным выжившим на этой стадии является желудевой червь, обладающий кровеносной системой с сердцем, которое также функционирует как почка. Желудевые черви имеют структуру, похожую на жабры, используемую для дыхания, структуру, аналогичную структуре примитивных рыб. У желудевых червей сплетение сосредоточено как в дорсальных, так и в вентральных нервных связках. Спинной канатик проникает в хоботок и частично отделяется от эпидермиса в этой области. Эта часть спинного нервного канатика часто является полой и вполне может быть гомологичной мозгу позвоночных. |
| Дата | Событие |
|---|---|
| 530 млн лет |  Pikaia Pikaia является культовым предком современных хордовых и позвоночных. Другие, более ранние хордовые предшественники включают Myllokunmingia fengjiaoa, Haikouella lanceolata и Haikouichthys ercaicunensis. Pikaia Pikaia является культовым предком современных хордовых и позвоночных. Другие, более ранние хордовые предшественники включают Myllokunmingia fengjiaoa, Haikouella lanceolata и Haikouichthys ercaicunensis. ланцетник, живущий сегодня, сохраняет некоторые характеристики примитивных хордовых. Он напоминает Pikaia. |
| 505 млн лет |  Агната Агната Появляются первые позвоночные : остракодермы, бесчелюстные рыбы, связанные с современными миногами и миксины. Haikouichthys и Myllokunmingia являются примерами этих бесчелюстных рыб, или Агнаты. (См. Также доисторическая рыба ). У них не было челюсти, а их внутренний скелет был хрящевым. У них отсутствовали парные (грудные и тазовые) плавники, как у более продвинутых рыб. Они были предшественниками Osteichthyes (костистая рыба). |
| 480 млн лет |  A плакодерма A плакодерма Плакодерми были доисторическими рыбами. Плакодермы были одними из первых челюстных рыб (Gnathostomata ), их челюсти произошли от первой жаберной дуги. Голова и грудная клетка плакодермы были покрыты сочлененными бронированными пластинами, а остальная часть тела была покрыта чешуей или обнажена. Однако летопись окаменелостей показывает, что они не оставили потомков после конца девона и менее связаны с живыми костными рыбами, чем акулы. |
| 410 млн лет назад |  Целакант пойман в 1974 г. Первый появляется латимерия ; этот отряд животных считался вымершим до тех пор, пока в 1938 году не были обнаружены живые экземпляры. Его часто называют живым ископаемым. Целакант пойман в 1974 г. Первый появляется латимерия ; этот отряд животных считался вымершим до тех пор, пока в 1938 году не были обнаружены живые экземпляры. Его часто называют живым ископаемым. |
| Дата | Событие |
|---|---|
| 390 млн лет |  Panderichthys Panderichthys Некоторые пресноводные рыбы с лепестковыми плавниками (Sarcopterygii ) развивают ноги и дают начало Tetrapoda. . Первые четвероногие эволюционировали в мелководных и болотистые пресноводные среды обитания. Примитивные четвероногие развились от рыб с лопастными плавниками («остеолепид саркоптериг »), с двудольный мозг в уплощенном черепе, широкий рот и короткая морда, чьи обращенные вверх глаза показывают, что это был обитатель дна, и который уже развил приспособления плавников с мясистым основанием и кости. («Живое ископаемое» латимерия - это родственная рыба с лопастными плавниками без этих приспособлений к мелководью.) Четвероногие рыбы использовали свои плавники в качестве лопастей в мелководных средах обитания, забитых растениями и растениями. детрит. Универсальные характеристики четвероногих - передние конечности, которые сгибаются назад в локтях, и задние конечности, которые сгибаются вперед в коленях, можно правдоподобно проследить до ранних четвероногих, живших на мелководье. Panderichthys - 90–130 см (35– 50 дюймов) длинная рыба из позднего девонского периода (380 Mya ). У него большая голова четвероногого. Panderichthys демонстрирует переходные черты между рыбами с лопастными плавниками и ранними четвероногими. Отпечатки путей, сделанные чем-то похожим на конечности Ихтиостеги, образовались 390 млн лет назад в польских морских приливных отложениях. Это говорит о том, что эволюция четвероногих древнее датированных окаменелостей Пандерихтиса до Ихтиостеги. Дыхательные рыбы сохраняют некоторые характеристики ранних тетрапод. Одним из примеров является двоякодышащая рыба Квинсленда. |
| 375 млн лет |  Тиктаалик Тиктаалик Тиктаалик - это род саркоптеригийских (лопастных) рыб из позднего девона с множеством четвероногих- как функции. Он показывает четкую связь между Пандерихтисом и Acanthostega. |
| 365 млн лет назад |  Acanthostega Acanthostega  Ichthyostega Ichthyostega Acanthostega - вымершим амфибией, среди первые животные с узнаваемыми конечностями. Это кандидат на звание одного из первых позвоночных, способных выходить на сушу. У него не было запястий, и в целом он был плохо приспособлен для жизни на суше. Конечности не выдерживали веса животного. Acanthostega имел и легкие, и жабры, что также указывало на то, что это была связь между рыбами с лопастными плавниками и наземными позвоночными. Ихтиостега - ранний четвероногий. Ichthyostega - одно из первых животных с ногами, руками и костями пальцев, и рассматривается как гибрид рыбы и земноводного. У Ихтиостеги были ноги, но их конечности, вероятно, не использовались для ходьбы. Они, возможно, проводили очень короткие периоды без воды и использовали бы свои ноги, чтобы прокладывать себе путь через грязь. Амфибии были первыми четвероногими животными, у которых развились легкие, которые, возможно, произошли от Хайнерпетон 360 млн лет назад. Земноводные, живущие сегодня, по-прежнему сохраняют многие характеристики ранних четвероногих. |
| 300 млн лет |  Hylonomus Hylonomus От земноводных произошли первые рептилии: Hylonomus - самый ранний из известных рептилия. Он был 20 см (8 дюймов) в длину (включая хвост) и, вероятно, был бы очень похож на современных ящериц. У него были маленькие острые зубы и, вероятно, он питался многоножками и ранними насекомыми. Это предшественник более поздних амниот и млекопитающих рептилий. Альфа-кератин первый развивается здесь. Он используется в когтях современных ящериц и птиц, а также в шерсти млекопитающих. Эволюция амниотического яйца дает начало амниотам, рептилиям, которые могут воспроизводиться на суше и откладывать яйца на суше. Им не нужно было возвращаться в воду для размножения. Эта адаптация дала им возможность впервые заселиться на возвышенностях. Рептилии обладают развитой нервной системой по сравнению с земноводными, с двенадцатью парами черепных нервов. |
| Дата | Событие |
|---|---|
| 256 Ма |  Phthinosuchus, ранний Therapsid Вскоре после появления первых рептилий отщепились две ветви. Одна ветвь - это Sauropsids, от которых произошли современные рептилии и птицы. Другая ветвь - Synapsida (Синапсиды), от которой произошли современные млекопитающие. У обоих были височные отверстия, пара отверстий в черепе за глазами, которые использовались для увеличения пространства для мышц челюсти. Синапсиды имели по одному отверстию с каждой стороны, тогда как диапсид (ответвление зауропсиды) имели два. Phthinosuchus, ранний Therapsid Вскоре после появления первых рептилий отщепились две ветви. Одна ветвь - это Sauropsids, от которых произошли современные рептилии и птицы. Другая ветвь - Synapsida (Синапсиды), от которой произошли современные млекопитающие. У обоих были височные отверстия, пара отверстий в черепе за глазами, которые использовались для увеличения пространства для мышц челюсти. Синапсиды имели по одному отверстию с каждой стороны, тогда как диапсид (ответвление зауропсиды) имели два. Самые ранние млекопитающие рептилии - это пеликозавры. Пеликозавры были первыми животными, у которых были височные оконные проемы. Пеликозавры - не терапсиды, но вскоре они их породили. Терапсиды были прямым предком млекопитающих. . У терапсидов височные оконные проемы больше и больше похожи на млекопитающих, чем пеликозавры, их зубы демонстрируют более последовательную дифференциацию, а более поздние формы развили вторичное небо. Вторичное нёбо позволяет животному есть и дышать одновременно и является признаком более активного, возможно, теплокровного образа жизни. |
| 220 млн. Лет |  Cynognathus Cynognathus Одна подгруппа терапсид, цинодонты приобрели более сходные с млекопитающими характеристики. Челюсти цинодонтов напоминают челюсти современных млекопитающих. Эта группа животных, вероятно, содержит вид, который является прямым предком всех современных млекопитающих. |
| 220 млн лет |  Repenomamus Repenomamus От Eucynodontia (cynodonts ) появились первые млекопитающие. Большинство ранних млекопитающих были маленькими землеройками, питавшимися насекомыми. Хотя в летописи окаменелостей нет доказательств, вполне вероятно, что у этих животных была постоянная температура тела и молочные железы для их детенышей. Область неокортекса мозга впервые возникла у млекопитающих и поэтому является уникальной для них. Одноножки - это откладывающая яйца группа млекопитающих, представленных среди современных животных утконосом и ехидной. Недавнее секвенирование генома утконоса показывает, что его половые гены ближе к генам птиц, чем к генам териан (живые роды) млекопитающих. Сравнивая это с другими млекопитающими, можно сделать вывод, что первые млекопитающие, получившие половую дифференциацию благодаря наличию или отсутствию гена SRY (обнаруженного в y-хромосоме), эволюционировали после монотремной родословная отделилась. |
| 160 млн лет назад |  Juramaia sinensis Juramaia sinensis Juramaia sinensis - самая ранняя из известных ископаемых млекопитающих. |
| 100 млн лет назад | Последний общий предок мышей и людей (основание клады Euarchontoglires ). |
| Дата | Событие |
|---|---|
| 85–66 млн лет |  Plesiadapis Plesiadapis  Carpolestes simpsoni Carpolestes simpsoni Группа мелких, ночных, древесных, насекомоядных млекопитающих Эуархонта начинает видообразование, которое приведет к отрядам приматов, древовидных и летающих лемуров. Primatomorpha - это подразделение Euarchonta, включая приматов и их предков-стволовых приматов Plesiadapiformes. Ранний примат-стебель, Plesiadapis, все еще имел когти и глаза по бокам головы, что делало его быстрее на земле, чем на деревьях, но он начал много времени проводить на нижних ветвях, питаясь плоды и листья. Plesiadapiformes, скорее всего, являются предками всех приматов. Впервые они появились в летописи окаменелостей около 66 миллионов лет назад, вскоре после вымирания мелового и палеогенового периода, которое уничтожило около трех четвертей видов растений и животных на Земле, включая большинство динозавров. Один из последних Plesiadapiformes - Carpolestes simpsoni, с хватательными пальцами, но не обращенными вперед глазами. |
| 63 млн лет | приматы делятся на подотряд Strepsirrhini (мокрые приматы) и Haplorrhini (сухоносые приматы). Strepsirrhini содержат большинство просимиан ; современные примеры включают лемуров и лорисов. Хаплоррины включают две живые группы: просимейские долгопяты и обезьяны обезьяны, включая обезьян. Одним из самых ранних haplorrhines является Teilhardina asiatica, дневное существо размером с мышь с маленькими глазами. Метаболизм Haplorrhini утратил способность производить витамин C, заставляя всех потомков включать в свой рацион фрукты, содержащие витамин C. |
| 30 млн лет назад |  Aegyptopithecus Aegyptopithecus Haplorrhini разделяется на инфраотряды Platyrrhini и Catarrhini. Platyrrhines, обезьяны Нового Света, имеют цепкий хвост, а самцы дальтоники. Предполагается, что люди, чьи потомки станут Platyrrhini, мигрировали в Южную Америку либо на плоту растительности, либо по сухопутному мосту (теперь гипотеза подтверждается). Катаррины в основном оставались в Африке, когда два континента отделились. Возможные ранние предки катарринов: Aegyptopithecus и Saadanius. |
| 25 млн лет назад |  Проконсул Проконсул Catarrhini разделены на 2 суперсемейства, обезьяны Старого Света (Cercopithecoidea) и обезьяны (Hominoidea ). Наше трехцветное цветовое зрение имело свои генетические корни в этот период. Проконсул был одним из первых представителей рода катаральных приматов. У них была смесь характеристик обезьяны Старого Света и обезьяны. Обезьяньи черты проконсула включают тонкую зубную эмаль, легкое телосложение с узкой грудью и короткими передними конечностями и древесный образ жизни на четвероногих. Его обезьяноподобные черты - отсутствие хвоста, обезьяноподобных локтей и немного больший мозг по сравнению с размером тела. Проконсул африканский - возможный предок как больших, так и малых обезьян, включая людей. |
| Дата | Событие |
|---|---|
| 18 млн лет | Hominidae (предки великих обезьян) происходят от предков гиббонов (малых обезьян) между c. 20–16 млн лет. |
| 16 млн лет | предки гоминин происходят от предков орангутанга между c. 18–14 млн лет. Pierolapithecus catalaunicus считается общим предком человека и других человекообразных обезьян или, по крайней мере, видом, который приближает нас к общему предку чем любое предыдущее открытие окаменелостей. У него были особые приспособления для лазания по деревьям, как у современных людей и других человекообразных обезьян: широкая плоская грудная клетка, жесткая нижняя часть позвоночника, гибкие запястья и лопатки, лежащие вдоль его спины. |
| 12 млн лет назад | Danuvius guggenmosi - первая обнаруженная в позднем миоцене большая обезьяна с сохранившимися длинными костями, которая в значительной степени объясняет анатомическое строение и передвижения современных обезьян. У него были приспособления как для висящего на деревьях (поддерживающее поведение ), так и для ходьбы на двух ногах (двуногие ), тогда как среди современных гоминидов люди лучше приспособлены к последним и другие для первого. Таким образом, у Данувиуса был метод передвижения, в отличие от любой ранее известной обезьяны, называемый «карабканье на вытянутых конечностях», он шел прямо по ветвям деревьев, а также использовал руки для подвешивания. Последний общий предок между людьми и другими обезьянами, возможно, имел аналогичный способ передвижения. |
| 10 млн. Лет | Клада, в настоящее время представленная людьми и родом Pan (обыкновенные шимпанзе и бонобо ), отделилась от предков горилл между c. От 10 до 8 млн лет. |
| 6 млн лет |  Sahelanthropus tchadensis Sahelanthropus tchadensis Hominini : последний общий предок людей и шимпанзе по оценкам, они жили примерно от 10 до 5 миллионов лет назад. И шимпанзе, и люди имеют гортань, которая перемещается в течение первых двух лет жизни в точку между глоткой и легкими, что указывает на то, что у общих предков есть эта особенность, что является предпосылкой для вокализированная речь у людей. Видообразование могло начаться вскоре после 10 млн лет назад, но позднее смешение между линиями могло произойти до 5 млн лет назад. Кандидаты в виды Hominina или Homininae, которые жили в этот период времени, включают Ouranopithecus (ок. 8 млн лет назад), Graecopithecus (ок. 7 Ma), Sahelanthropus tchadensis (около 7 млн лет), Orrorin tugenensis (около 6 млн лет). 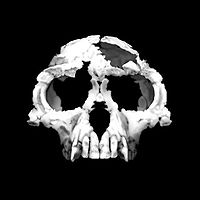 Ardipithecus Ardipithecus является или может быть очень ранним hominin родом (tribe Hominini и подтриба Гоминина ). В литературе описаны два вида: A. ramidus, который жил около 4,4 миллиона лет назад в начале плиоцена, и A. kadabba, датируемый примерно 5,6 миллиона лет назад (поздний миоцен ). A. ramidus имел небольшой мозг размером от 300 до 350 см. Он примерно такого же размера, как мозг современного бонобо и самки обыкновенного шимпанзе ; он несколько меньше, чем мозг австралопитеков, таких как Люси (от 400 до 550 см), и немногим более одной пятой размера мозга современного Homo sapiens. Ardipithecus Ardipithecus является или может быть очень ранним hominin родом (tribe Hominini и подтриба Гоминина ). В литературе описаны два вида: A. ramidus, который жил около 4,4 миллиона лет назад в начале плиоцена, и A. kadabba, датируемый примерно 5,6 миллиона лет назад (поздний миоцен ). A. ramidus имел небольшой мозг размером от 300 до 350 см. Он примерно такого же размера, как мозг современного бонобо и самки обыкновенного шимпанзе ; он несколько меньше, чем мозг австралопитеков, таких как Люси (от 400 до 550 см), и немногим более одной пятой размера мозга современного Homo sapiens. Ардипитек был древесным, что означает, что он жил в основном в лесу, где он конкурировал с другими лесными животными за пищу, без сомнения, включая современного предка шимпанзе. Ардипитек, вероятно, был двуногим, о чем свидетельствуют его чашеобразный таз, угол большого затылочного отверстия и более тонкие кости запястья, хотя его ступни все еще были приспособлены для хватания, а не для ходьбы на большие расстояния. |
| 3,6 млн. Лет | Член Australopithecus afarensis оставил человеческие следы на вулканическом пепле в Лаэтоли, северная Танзания, что дает убедительные доказательства того, что полный рабочий день прямохождение. Australopithecus afarensis жил между 3,9 и 2,9 миллиона лет назад и считается одним из самых ранних гомининов - тех видов, которые развились и составили линию Homo и ближайших родственников Homo после отделения от линии шимпанзе. Считается, что A. afarensis был предком как рода Australopithecus, так и рода Homo. По сравнению с современными и вымершими человекообразными обезьянами у A. afarensis были уменьшены клыки и коренные зубы, хотя они все еще были относительно крупнее, чем у современного человека. A. afarensis также имеет относительно небольшой размер мозга (380–430 см³) и прогнатическое (выступающее вперед) лицо. Австралопитеки были обнаружены в среде саванн; они, вероятно, разработали свою диету, чтобы включить в нее мясо. Анализ Australopithecus africanus нижних позвонков позволяет предположить, что эти кости изменились у женщин для поддержки двуногости даже во время беременности. |
| 3,5–3,3 млн лет. | Kenyanthropus platyops, возможный предок Homo, произошел от австралопитека. Каменные орудия труда изготовлены намеренно. |
| 3 млн. Лет | Двуногие австралопитеки (род подтрибы Hominina ) развиваются в саваннах Африки, за которым охотится Мегантереон. Выпадение волос на теле происходит от 3 до 2 млн лет назад, параллельно с развитием полного двуногого мышления. |
| Дата | Событие |
|---|---|
| 2,5–2,0 млн лет назад | Ранний Homo появляется в Восточной Африке, происходя от австралопитеков предков. Сложные каменные орудия знаменуют начало нижнего палеолита. Australopithecus garhi использовал каменные орудия примерно 2,5 млн лет назад. Homo habilis - самый старый вид, получивший обозначение Homo по Лики и др. (1964). H. habilis занимает промежуточное положение между Australopithecus afarensis и H. erectus, и были предложения отнести его к роду Australopithecus как Australopithecus habilis. Каменные орудия, найденные на стоянке Шангчен в Китае и датированные 2,12 миллиона лет назад, считаются самыми ранними известными свидетельствами существования гомининов за пределами Африки, превосходящими Дманиси в Грузии на 300 000 лет. |
| 1,9–0,5 млн лет | Homo erectus происходит от раннего Homo или позднего австралопитека. Homo habilis, хотя и значительно отличается по анатомии и физиологии, считается предком Homo ergaster, или африканского Homo erectus; но также известно, что он сосуществовал с H. erectus почти полмиллиона лет (примерно до 1,5 млн лет назад). С момента своего появления примерно 1,9 млн лет назад H. erectus распространен в Восточной Африке и Юго-Западной Азии (Homo georgicus ). H. erectus - первый известный вид, который научился контролировать огонь примерно на 1,5 млн лет. H. erectus позднее мигрирует по Евразии, достигнув Юго-Восточной Азии на 0,7 млн лет. Он описан в ряде подвидов. . Эволюция темной кожи примерно 1,2 млн лет. Homo antecessor может быть общим предком людей и неандертальцев. По нынешним оценкам, люди имеют примерно 20 000–25 000 генов и разделяют 99% своей ДНК с ныне вымершими неандертальцами и 95–99% их ДНК с их ближайшим живым эволюционным родственником, шимпанзе. Было обнаружено, что человеческий вариант гена FOXP2 (связанный с контролем речи) идентичен у неандертальцев. |
| 0,8–0,3 млн лет | Дивергенция неандертальцев и Денисовские линии от общего предка. Homo heidelbergensis (в Африке также известный как Homo rhodesiensis ) долгое время считался вероятным кандидатом на роль последнего общего предка неандертальцев и современных людей. Однако генетические данные из окаменелостей Sima de los Huesos, опубликованные в 2016 году, по-видимому, предполагают, что H. heidelbergensis в целом следует включить в линию неандертальцев как «до-неандертальцев» или «ранних неандертальцев». в то время как время расхождения между неандертальскими и современными линиями было отодвинуто назад до появления H. heidelbergensis, примерно от 600 000 до 800 000 лет назад, приблизительный возраст Homo antecessor. Затвердевшие следы датируются около 350 тыс. лет назад и связанные с H. heidelbergensis были обнаружены на юге Италии в 2003 г. |
| Дата | Событие |
|---|---|
| 300–130 тыс. лет назад |  Реконструкция H. neanderthalensis Реконструкция H. neanderthalensis Окаменелости, приписываемые H. sapiens, а также каменные орудия, датируемые примерно 300 000 лет назад, найденные в Джебель-Ирхуд, Марокко, дают самые ранние ископаемые свидетельства анатомически современного Homo sapiens. Современное человеческое присутствие в Восточной Африке (Гадемотта ), 276 тыс. Лет назад. Окаменелость челюстной кости возрастом 177000 лет, обнаруженная в Израиле в 2017 году, является старейшим человеческим останком, найденным за пределами Африки. Однако в июле 2019 года антропологи сообщили об обнаружении 210 000-летних останков H. sapiens и 170 000-летних останков H. neanderthalensis в пещере Апидима, Пелопоннес, Греция, более чем на 150 000 лет старше, чем предыдущие находки H. sapiens в Европе. Неандертальцы произошли от линии Homo heidelbergensis примерно в то же время (300 тыс. Лет назад). Патрилинейный и матрилинейный самые недавние общие предки (MRCA) живых людей примерно от 200 до 100 тыс. Лет назад, при этом некоторые оценки патрилинейного MRCA несколько выше - от 250 до 500 тыс. Лет назад. 160000 лет назад Homo sapiens idaltu в долине реки Аваш (недалеко от современной деревни Херто, Эфиопия) практиковал разрушение. |
| 130–80 тыс. Лет назад | морской изотоп 5-й этап (эемский ). Современное человеческое присутствие в Южной Африке и Западной Африке. Появление митохондриальной гаплогруппы (mt-haplogroup) L2. |
| 80–50 тыс. Лет | МИС 4, начало верхнего палеолита. Ранние свидетельства модерна поведения. Появление мт-гаплогрупп M и N. Южное расселение миграция из Африки, прото-австралоиды заселение Океании. примесь архаики от неандертальцев в Евразии, от денисовцев в Океании со следовыми количествами в Восточной Евразии, а также от неустановленной африканской линии архаичных людей в южной части Сахары Африка, а также скрещенные виды неандертальцев и денисовцев в Азии и Океании. |
| 50–25 тыс. Лет назад |  Реконструкция Оазе 2 (около 40 тыс. Лет назад) Реконструкция Оазе 2 (около 40 тыс. Лет назад) Поведенческая современность развивается в соответствии с теорией "большого скачка вперед". Исчезновение мутации Homo floresiensis. M168 (переносится всеми мужчинами неафриканского происхождения). Появление мт-гаплогрупп U и K. Население Европы, население Северной Азии Мамонтовой степи. Искусство палеолита. Вымирание неандертальцев и других архаичных человеческих вариантов (с возможным выживанием гибридных популяций в Азии и Африке.) Появление Y-гаплогруппы R2 ; mt-гаплогруппы J и X. |
| после 25 тыс. лет назад | Последний ледниковый максимум ; Эпипалеолит / мезолит / голоцен. Население Америки. Внешний вид: Y-гаплогруппа R1a ; mt-гаплогруппы V и T. Различные недавние расхождения, связанные с давлением окружающей среды, например светлая кожа у европейцев и восточных азиатов (KITLG, ASIP ), после 30 тыс. лет назад; инуитов адаптация к диета с высоким содержанием жиров и холодный климат, 20 тыс. лет назад Вымирание поздних выживших архаичных людей в начале голоцена (12 тыс. лет назад). Ускоренная дивергенция из-за давления отбора в популяциях, участвовавших в неолитической революции после 12 тыс. Лет назад, например Восточноазиатские типы ADH1B, связанные с одомашниванием риса или стойкостью лактазы. |