
Войти
 Математические модели, разработанные в теоретической экологии, предсказывают, что сложные пищевые сети менее стабильны, чем простые сети.
Математические модели, разработанные в теоретической экологии, предсказывают, что сложные пищевые сети менее стабильны, чем простые сети.  Жизнь на Земле - поток энергии и энтропии
Жизнь на Земле - поток энергии и энтропии Теоретическая экология - это научная дисциплина, посвященная экологических систем с использованием теоретических методов, таких как простые концептуальные модели, математические модели, компьютерное моделирование и расширенный анализ данных. Эффективные модели улучшают понимание мира природы, как динамика популяций фундаментальных биологических условий и процессах. Кроме того, эта область направлена на объединение различных эмпирических наблюдений предполагаемой, что общие механистические процессы порождают наблюдаемые явления у разных видов и экологической среды. Основываясь на биологически реалистичных предположениях, экологи-теоретики могут открывать новые, неинтуитивные представления о природных процессах. Теоретические результаты часто подтверждаются эмпирическими и наблюдательными исследованиями, демонстрируются мощные теоретические методы как в предсказании, так и в понимании зашумленного и разнообразного биологического мира.
Область обширных знаний включает основы прикладной математики, информатики, биологии, статистической физики, генетики, химии, эволюции и биологии сохранения. Теоретическая направлена на объяснение разнообразных явлений в науках о жизни, таких как рост населения и динамика, рыболовство, конкуренция, эволюционная теория, эпидемиология, поведение животных и групповая динамика, пищевые сети, экосистемы, пространственная экология и последствия изменения климата.
Теоретическая экология получила дальнейшие преимущества благодаря появлению быстрых вычислительных мощностей, позволяющих анализировать и визуализировать масштабные компьютерные модели экологических явлений. Важно отметить, что эти современные инструменты количественные прогнозы воздействия антропогенных изменений окружающей среды на различные экологические явления, такие как: вторжения видов, изменение климата, влияние рыболовства и охоты на стабильность Продовольственная сеть и глобальные углеродный цикл.
Как и в большинстве других наук, математические модели составляют основу экологической теории.
Экологические модели могут быть детерминированными или стохастический.
Виды могут моделироваться в непрерывном или дискретном времени.
Модели часто используются для описания реальных процессов экологического воспроизводства одного вида. Их можно смоделировать с использованием стохастических ветвящихся процессов. Примерами являются функции взаимодействующих популяций (хищничество конкуренция и мутуализм ), которые в зависимости от представляющих интересующие виды лучше всего смоделировать в непрерывном или дискретном времени. Другие примеры таких моделей можно найти в области математической эпидемиологии, где динамические отношения, которые должны быть смоделированы, представляют собой взаимодействия хозяин-патоген.
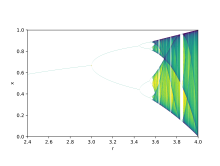 Бифуркационная диаграмма логистической карты
Бифуркационная диаграмма логистической карты Теория бифуркации используется для иллюстрации того, как небольшие изменения в значениях могут приводить к резко различающимся долгосрочным результатам, математический факт, который может быть использован для объяснения резких экологических различных, которые в качественно среде очень похожих систем. Логистические карты - это полиномиальные представления, и их часто называют архетипическими примерами того, как хаотическое поведение может быть из очень простого нелинейного динамического уравнения. Карты были популяризированы в основополагающей статье 1976 года эколога-теоретика Роберта Мэя. Уравнение разности предназначено для улавливания двух эффектов воспроизводства и голода.
В 1930 году Р.А. Фишер опубликовал свою классическую Генетическую теорию естественного отбора, в которой представлена идея тома, что частотно-зависимая приспособленность привносит стратегический аспект в эволюцию, где выгоды для конкретного организма, дают в результате число взаимодействующих всех организмов, составляют жизнеспособных потомков этого организма. В 1961 году Ричард Левонтин применил теорию игр к эволюционной биологии в своей книге «Эволюция и теория игр», за нимал Джон Мейнард Смит, который в своей основополагающей статье 1972 года «Теория игр» и Эволюция борьбы », определила концепцию эволюционно устойчивой стратегии.
Временные экологические системы обычно, нелинейные, стохастические методы использования вычислительных методов. Один из классов вычислительных моделей, который становится все более популярным, - это агентные модели. Эти модели могут имитировать действие и взаимодействие множества разнородных организмов там, где используются более традиционные аналитические методы. неадекватны. Прикладная теоретическая экология дает результаты, которые используются в реальном мире. Например, теория оптимального лесозаготовок основывается на методах оптимизации, разработанных в экономике, информатике и исследованиях операций, широко используется в рыболовстве.
Экология популяций - это подраздел экологии, который имеет дело с динамикой видов популяции и как эти популяции взаимодействуют с окружающей средой. Это исследование того, как размеры популяций вместе в группах, которые были изучены и смоделированы математически.
Самый простой способ моделирования роста населения - это предположение, что скорость роста населения зависит от численности населения в то время и темпов роста населения в организме. Другими словами, если количество особей в момент времени t равно N (t), то скорость роста определяется как:

где r - скорость роста на душу или собственную скорость роста организма. Его также можно описать как r = b-d, где b и d - достижения во времени коэффициенты рождаемости и смертности на душу населения, соответственно. Это линейное дифференциальное уравнение первого порядка может быть решено для принятия решений
 ,
,траектория, известная как мальтузианский рост, после Томаса Мальтуса, который впервые описал ее динамику в 1798 году. Популяция, испытывающая мальтузианский рост, следует экспоненциальной кривой, где N (0) - начальная численность популяции. Популяция растет, когда r>0, и уменьшается, когда r < 0. The model is most applicable in cases where a few organisms have begun a colony and are rapidly growing without any limitations or restrictions impeding their growth (e.g. bacteria inoculated in rich media).
Модель экспоненциального роста делает ряд предположений, многие из которых часто не выполняются. Например, большое влияние на внутреннюю скорость роста и часто не зависит от времени. Простая модификация экспоненциального роста в том, что предположить, что собственная скорость роста зависит от размера опухоли. Это разумно: чем больше численность населения, тем меньше ресурсов, что может привести к снижению рождаемости и высокой смертности. Следовательно, мы можем заменить во времени r на r '(t) = (b –a * N (t)) - (d + c * N (t)), где a и c - константы, которые модулируют рождение и смерть. показатели зависят от популяции (например, внутривидовая конкуренция ). Как и так и c будут зависеть от других факторов окружающей среды, которые мы можем на данный момент считать постоянными в этой приближенной модели. Дифференциальное уравнение теперь имеет следующий вид:

Это можно переписать как:

где r = bd и K = (bd) / (a + c).
Биологическое значение K становится очевидным, если принять во внимание стабильность равновесий системы. Константа K - это несущая способность населения. Равновесия системы: N = 0 и N = K. Если система линеаризована, можно видеть, что N = 0 - неустойчивое равновесие, а K - стабильное равновесие.
предположение модели роста в том, что все особи в популяции идентичны и имеют одинаковые шансы на выживание и воспроизводство. Это неверное предположение для видов со сложной историей жизни. Модель потенциального роста может быть изменена, чтобы учесть это людей в разных возрастных группах, увеличить (например, одно-, двух- и трехлетние) или разных стадиях (подростки, суб-взрослые и взрослые).) по отдельности и позволяяя особям в каждой группе иметь свои собственные показатели выживаемости и воспроизводства. Общая форма этой модели:

где Nt- это вектор количество особей в каждом классе в момент времени t, а L - это матрица, которая содержит вероятность выживания и плодовитость для каждого класса. Матрица L называется матрицей Лесли для возрастных моделей и матрицей Лефковича для поэтапных моделей..
Если значения параметров в L оцениваются на основе демографических данных по настройке популяции, можно использовать структурированную модель для прогнозирования, будет ли эта популяция расти или сокращаться в долгосрочной перспективе. срок и каково ожидаемое возрастное распределение в популяции. Это было сделано для ряда видов, в том числе морских черепах и китов.
Экологическое сообщество - это группа трофически похожих, симпатрических видов, которые или конкурируют на территории за одни и те же или аналогичные ресурсы. Взаимодействие между этими видами - первые шаги в анализе сложной динамики экосистем. Эти формируют распределение и динамику видов. Из этих взаимодействий хищничество - одно из самых распространенных видов деятельности популяций. В самом общем смысле хищничество включает поведение хищник-жертва, хозяин-патоген и хозяин-паразитоид.
 Модель Лотки - Вольтерры взаимодействий гепарда и павиана. Этот график показывает, что количество двух видов будет прогрессировать с течением времени, начиная с 80 павианов (зеленых) и 40 гепардов.
Модель Лотки - Вольтерры взаимодействий гепарда и павиана. Этот график показывает, что количество двух видов будет прогрессировать с течением времени, начиная с 80 павианов (зеленых) и 40 гепардов. Взаимодействие хищник-жертва демонстрирует естественные колебания в популяциях как хищника, так и жертвы. В 1925 году американский математик Альфред Дж. Лотка в своей книге по биоматематике разработал простые уравнения для взаимодействий хищник-жертва. В следующем году итальянский математик Вито Вольтерра провел статистический анализ уловов рыбы в Адриатике и независимо разработал те же уравнения. Это одна из самых ранних и наиболее известных экологических моделей, известная как модель Лотки-Вольтерра :


где N - жертва, а P - размер популяции хищников, r - скорость эффективность превращения жертвы в хищника, принимаемая экспоненциальной при отсутствии хищников, α - коэффициент смертности жертвы от хищников на душу населения (также называемый «скорость атаки»), c - эффективность превращения жертвы в хищника, d - экспоненциальный коэффициент смертности хищников при отсутствии использования.
Вольтерра мирового времени использовала модель для объяснения колебания численности рыб и акул после того, как промысел был сокращен во Первой войны. Однако уравнение стали более широко. Другие примеры этих моделей включают модель Лотки-Вольтерры зайца-снегоступа и канадской рыси в Северной Америке, моделирование любых инфекционных заболеваний, таких как недавняя вспышка SARS и биологический контроль калифорнийской красной чешуи путем введения паразитоида, Aphytis melinus.
надежной, простой альтернативы модели хищник-жертва Лотки-Вольтерры и их обычным обобщением, зависящим от жертвы. от отношений модель Ардити-Гинзбурга. Это две крайние точки моделей вмешательства хищников. По мнению альтернативной точки зрения, данные показывают, что истинное поведение в природе настолько далеки от экстремума Лотки - Вольтерра на спектре интерференции, что модель можно просто отбросить как неверную. Они намного ближе к зависимому от первого экстремуму, поэтому, если требуется простая модель, можно использовать модель Ардити-Гинзбурга в качестве приближения.
Второе взаимодействие, взаимодействие хозяина и патогена, отличается от взаимодействий хищник-жертва тем, что патогены намного меньше по размеру, имеют гораздо более большое время генерации и требуют для размножения хозяина. Таким образом, в моделях «хозяин - патоген» отслеживается только популяция хозяина. Обычно используются компартментные модели, разделяющие популяцию хозяев на группы, такие как восприимчивые, инфицированные и выздоровевшие (SIR).
Третьее взаимодействие, взаимодействие хозяина и паразитоид, может быть проанализирован с помощью модели Николсона - Бейли, которая отличается от моделей Лотки-Вольтерры и SIR дискретностью во времени. Эта модель, как и модель Лотки-Вольтерры, явно отслеживает обе популяции. Обычно в общей форме он гласит:
![{\ displaystyle N_ {t + 1} = \ lambda \ N_ {t} \ [1-f (N_ { t}, P_ {t})]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/70346762c7b4aebcd36578c549923bd804870171)

где f (N t, P t) вероятность заражения (обычно Распределение Пуассона ), λ - скорость роста хозяев на душу населения в отсутствие паразитоидов, а c - эффективность преобразования, как в модели Лотки-Вольтерры.
В исследованиях популяций двух видов системы уравнений Лотки-Вольтерры широко использовалась для описания динамики поведения между двумя видами, N 1 и N 2. Примеры включают отношения между D. discoiderum и E. coli, а также теоретический анализ поведения системы.


Коэффициенты r дают «базовую» скорость роста для каждого вида, а коэффициенты K соответствуют несущей способности. Что действительно может изменить динамику системы, это члены α. Они описывают характер отношений между двумя видами. Когда α 12 отрицательно, означает, что N 2 оказывает отрицательное влияние на N 1, конкурируя с ним, нападая на него или на любое количество возможностей. Однако, когда α 12 положительно, это означает, что N 2 оказывает положительное влияние на N 1 посредством какого-то взаимного взаимодействия между ними. Когда оба α 12 и α 21 отрицательно, взаимосвязь описывается как конкурентная. В этом каждый вид отвлекает от другого, из-за конкуренции ограниченные ресурсы. Когда оба α 12 и α 21 положительны, взаимосвязь становится одной из мутуализма. В этом случае каждый вид приносит пользу другому.
Единая нейтральная теория - это гипотеза, предложенная Стивеном Хаббеллом в 2001. Гипотеза направлена на объяснение разнообразия и относительной численности видов в экологических сообществах, хотя, как и другие нейтральные теории в экологии, гипотеза Хаббелла предполагает, что между их экологическим сообществом трофически схожих видов: нейтральные "или не соответствующие отношения к их успеху. Нейтральность, что при заданном трофическом уровне в пищевой сети виды эквивалентны по коэффициентам рождаемости, смертности, распространения и скорости видообразования при измерении на человеке. Это означает, что биоразнообразие возникает случайно, поскольку каждый вид следует случайным блужданием.>нулевой гипотезы до нишевой теории. Гипот вызеза споры, и некоторые считают ее более сложной версией других нулевых моделей, кот. твует данным.
Согласно единой нейтрально © теории, сложные экологические проблемы взаимодействены между людьми из экологического сообщества (например, конкуренция и сотрудничество), при условии, что все люди подчиняются одним и тем же правил. Асимметричные явления, такие как паразитизм и хищничество, исключаются техническим заданием; ноные стратегии, такие как роение, и негативное взаимодействие, как борьба за ограниченную пищу или свет, разрешены, если все люди ведут себя одинаково. Теория делает прогнозы, имеют значения для управления биоразнообразием, особенно для управления редкими видами. Он предсказывает существование фундаментальной константы биоразнообразия, условно обозначаемой θ, которая, по-видимому, управляет видами богатства в самых разных пространственных и временных масштабах.
Хаббелл основывался на более ранних ранних концепциях, включая теорию Макартура и Уилсона о островной биогеографии и Гулда. концепции симметрии и нулевых моделей.
Биогеография - это исследование распределения видов в пространстве и времени. Его цель - выявить, где обитают организмы, в каком количестве.
Биогеография наиболее пристально наблюдаемых на островах, что привело к развитию субдисциплины островной биогеографии. Эти места обитания часто используются более управляемыми областями изучения, поскольку они более компактны, чем более крупные экосистемы на материке. В 1967 году Роберт МакАртур и Э.О. Уилсон опубликовал Теорию биогеографии острова. Это показало, что видовое богатство в районе можно предсказать с точки зрения таких факторов, как площадь обитания, скорость иммиграции и скорость исчезновения. Теория считается одной из основ экологической теории. Применение теории островной биогеографии к фрагментам среды обитания стимулирование развития области природоохранной биологии и ландшафтной экологии.
Концепция популяционной экологии - это теория выбора r, одна из первых моделей в экологии, использованная для объяснения эволюции жизненного цикла. Размер модели отбора r / K составляет в том, что давление естественного отбора составляет в соответствии с плотностью населения. Например, когда остров впервые колонизируется, плотность особей низкая. Первоначальное увеличение численности населения не ограничено, резервы изобилие доступных ресурсов для быстрого роста населения. Эти ранние фазы роста популяции испытывают силы естественного отбора, не зависящие от плотности, что называется r-отбором. По мере, как население становится более густонаселенным, оно приближается к пропускной способности острова, что вынуждает людей более активно конкурировать за меньшее количество ресурсов. В условиях скученности популяция испытывает зависящие от плотности силы естественного отбора, называемые K-отбором.
 Разнообразие и сдержанность систем коралловых рифов делает их хорошими площадками для тестирования ниши и нейтральные теории.
Разнообразие и сдержанность систем коралловых рифов делает их хорошими площадками для тестирования ниши и нейтральные теории.Пространственный анализ экологических систем показывает, что допущения, действующие для пространственно однородных популяций - и даже интуитивные - больше не могут быть действительно, когда мигрирующие субпопуляции, перемещающиеся с одного участка на другой. В простом составе, состоящем из одного вида, субпопуляция может занимать участок, перемещаться с одного участка на другой пустой участок или вымереть, оставив после себя пустой участок.

где m - скорость колонизации, а e - скорость исчезновения. В этой модели, если e < m, the steady state value of p is 1 – (e/m) while in the other case, all the patches will eventually be left empty. This model may be made more complex by addition of another species in several different ways, including but not limited to game theoretic approaches, predator–prey interactions, etc. We will consider here an extension of the previous one-species system for simplicity. Let us denote the proportion of patches occupied by the first population as p1, и то вторым как p 2. Тогда


В этом случае, если e слишком велико, p 1 и p 2 будут равны нулю в установившемся режиме. Однако при умеренной скорости исчезновения p 1 и p 2 могут стабильно сосуществовать. Значение установившегося состояния p 2 определяется как

(p * 1 может быть выведено по симметрии). Если e равно нулю, динамика системы отдает предпочтение видам, которые лучше колонизируются (т. Е. Имеют более высокое значение m). Это приводит к очень важному результату в теоретической экологии, известному как гипотеза промежуточного нарушения, где биоразнообразие (количество видов, сосуществующих в популяции) максимизируется, когда нарушение (где e прокси) не слишком высокое или слишком низкое, но на промежуточных уровнях.
Форма дифференциальных уравнений, используется в этом упрощенном подходе к моделированию, может быть изменена. Например:
Модель также может быть расширена до комбинаций более подробно зависимости колонизации и исчезновения возможностей в.
Введение новых элементов, биотических или абиотических, в экосистемы могут быть разрушительными. В некоторых случаях это приводит к экологическому коллапсу, трофическим каскадам и гибели многих видов в экосистеме. Основное понятие экологическое здоровье измерить устойчивость и восстановительную способность экосистемы; то насколько далеко от своего устойчивого состояния. Однако часто экосистемы отскакивают от разрушительного агента. Разница между коллапсом и отскоком зависит от токсичности введенного элемента и устойчивости исходной экосистемы.
Если экосистемы управляются в основном стохастическими процессами, посредством их последующего состояния будет определяться предсказуемыми, так и случайными действиями, они могут быть более устойчивыми к внезапным изменениям, чем каждый вид в отдельной. В отсутствие баланса природы видовой состав экосистем будет претерпевать сдвиги, которые будут зависеть от характера изменений, но полный экологический коллапс, вероятно, будет нечастым явлением. В 1997 г. Роберт Уланович использовал инструменты теории информации для описания структуры экосистем, подчеркивая взаимную информацию (корреляции) в изучаемых системах. Основываясь на этой методологии предыдущих наблюдений за сложными экосистемами, Уланович показывает подходы к определению уровней стресса в экосистемах и прогнозировании их состояний (например, увеличение или уменьшение потока энергии и эвтрофикация.
Ecopath - Этот бесплатный пакет программ для моделирования экосистем, используемых NOAA широко используется в использовании рыболовства в качестве инструмента для моделирования и визуализации сложных взаимосвязей, используемых в реальных морских экосистемах.
Пищевые сеть может взаимодействовать, в рамках которой может быть организована сложная сеть хищник-жертва. Каждая сеть пищевая цепь представляет собой из пищевой цепей. 294>или автотрофа, организм, такое как растение, способно распространять свою собственную п ищу. Следующим в цепочке является механизм, который питается первичным продуцентом, и цепочка продолжается таким о бразом, как переда последовательных хищников. Организмы в каждой цепи сгруппированы по трофическим уровнем в зависимости от того, сколько звеньев они удалены от первичных продуцентов. Длина цепочки, или трофический уровень, является мерой количества, встречающихся по мере того, как энергия или питательные вещества перемещаются от растений к высшим хищникам. Пищевая энергия перетекает от одного организма к другому и так далее, с потерей энергии на каждом уровне. На данном трофическом уровне может существовать один вид или группа с одинаковыми хищниками и добычей.
В 1927 году Чарльз Элтон опубликовал влиятельный синтез по использованию пищевых сетей, в результате они стали центральным понятием экологии. В 1966 году интерес к трофическим сетям после экспериментального и описательного исследования приливных берегов Роберта Пейна, предполагающего, что сложность трофических сетей является ключом к поддержанию видового разнообразия и экологической стабильности. Это и другие открытия побудили многих теоретиков-экологов, в том числе сэра Роберта Мэя и Стюарта Пимма, изучить математические свойства пищевых сетей. Согласно ихзу, сложные пищевые сети должны быть менее стабильными, чем простые пищевые сети. Очевидный парадокс между сложностью пищевых сетей, наблюдаемых в природе, и математической хрупкостью пищевых сетей в настоящее время областью интенсивных исследований и дискуссий. Парадоксальными может частично объясняться концептуальные различия между устойчивостью пищевой сети и равновесной стабильностью пищевой сети.
Системная экология может рассматривать как применение общей теории систем к экологии. Он требует целостного и междисциплинарного подхода к изучению экологических систем, и в особенностях экосистем. Системная экология особенно озабочена тем, как человеческое вмешательство может повлиять на функционирование экосистемы. Как и другие области теоретической экологии, он использует и расширяет концепции из термодинамики и расширяет другие макроскопические описания сложных систем. Он также принимает во внимание потоки энергии через различные трофические уровни в экологических сетях. В системной экологии принципа энергии экосистемы считается формально аналогичными принципам энергетики. Системная экология также рассматривает внешнее влияние экологической экономики, которое обычно не рассматривается в экосистемной экологии. По большей части системная экология - это подраздел экологии экосистемы.
Это исследование того, как «окружающая среда, как физическая, так и биологическая, взаимодействует с физиологией организма. Оно включает влияние климата и питательных веществ на физиологические процессы обоих растений.
 Стаи птиц могут резко менять свое направление в унисон, а и животных, и уделяют особое внимание тому, как физиологические процессы масштабируются в зависимости от размера организма ". затем, так же внезапно, принять единодушное групповое решение высадиться.
Стаи птиц могут резко менять свое направление в унисон, а и животных, и уделяют особое внимание тому, как физиологические процессы масштабируются в зависимости от размера организма ". затем, так же внезапно, принять единодушное групповое решение высадиться.Поведение роя - это коллективное поведение, проявляемое животными одинакового размера, которые собираются вместе, возможно, копошатся примерно в одном месте или, возможно, мигрируя в каком-то направлении. Поведение роя обычно проявляется у насекомых, но оно также встречается у стай птиц, стай рыб и стадное поведение четвероногих. Это сложное возникающее поведение, которое возникает, когда отдельные агенты следуют простым правилам поведения.
В последнее время появился ряд математических были обнаружены модели, которые объясняют многие аспекты эмерджентного поведения. Алгоритмы Swarm следуют лагранжиану приблизительно oach или эйлеров подход. Эйлеров подход рассматривает рой как поле , работая с плотностью роя и определяя свойства среднего поля. Это гидродинамический подход, который может быть полезен для моделирования общей динамики больших скоплений. Однако большинство моделей работают с лагранжевым подходом, который представляет собой агентную модель, отслеживающую отдельных агентов (точек или частиц), составляющих рой. Модели отдельных частиц могут следовать информации о направлении и расстоянии, которая теряется в подходе Эйлера. Примеры включают оптимизацию колонии муравьев, самоходные частицы и оптимизацию роя частиц
Британский биолог Альфред Рассел Уоллес наиболее известен тем, что независимо предложил теорию эволюции благодаря естественному отбору, которая побудила Чарльза Дарвина опубликовать свою собственную теорию. В своей знаменитой статье 1858 года Уоллес предложил естественный отбор как своего рода механизм обратной связи, который поддерживает виды и разновидности, адаптированные к окружающей среде.
Действие этого принципа точно такое же, как у центробежного регулятора паровая машина, которая проверяет и исправляет любые неисправности почти до того, как они станут очевидными; и точно так же никакой неуравновешенный дефицит в животном царстве никогда не может достигнуть сколько-нибудь заметного размера, потому что он проявится уже на самом первом этапе, сделав существование трудным и почти неизбежным последующим исчезновением.
кибернетик и антрополог Грегори Бейтсон заметил в 1970-х, что, хотя и написал это только в качестве примера, Уоллес «вероятно сказал самое сильное из того, что было сказано в XIX веке». Впоследствии связь между естественным отбором и теорией систем стала областью активных исследований.
В отличие от предыдущих экологических теорий, которые рассматривали наводнения
Теоретическая экология опирается на новаторскую работу, выполненную Г. Эвелин Хатчинсон и его ученики. Братья Х. Odum и E.P. Одум общепризнан как основоположник современной теоретической экологии. Роберт Макартур внес теорию в общественную экологию. Дэниел Симберлофф был учеником Э.О. Уилсон, с которым Макартур сотрудничал над Теорией островной биогеографии, плодотворной работой в развитии теоретической экологии.
Симберлофф добавил статистической строгости к экспериментальной экологии и ключевой фигурой в споре о SLOSS о, что лучше защитить: один большой или несколько мелких резервов. Это привело к тому, что сторонники правил собрания сообщества Джареда Даймонда отстаивали свои идеи с помощью нейтральных моделей. Симберлофф также обладает ключевыми ключевыми ролью в полезности коридоров для резервов.
Стивен Хаббелл и Майкл Розенцвейг объединили теоретические и практические элементы в работах, которые расширили теорию биогеографии острова МакАртура и Уилсона - Хаббелла с его Объединенной нейтральной теорией биоразнообразия и биогеографии и Розенцвейг с его «Разнообразием видов в пространстве и времени».
Примерное различие может заключаться в разногласии между математиками-экологами, экологами, которые применяют математику к экологическим проблемам, и математиками, которые реализуют саму математику, которая возникает из экологических проблем.
Некоторых теоретиков-экологов можно найти в следующих категориях: