
Войти
| Вкусовые рецепторы | |
|---|---|
 Вкусовые рецепторы языка присутствуют во вкусовых сосочках сосочков. Вкусовые рецепторы языка присутствуют во вкусовых сосочках сосочков. | |
| Идентификаторы | |
| FMA | 84662 |
| Анатомическая терминология [редакция Викиданных ] | |
A вкусовый рецептор - это тип рецептора, который способствует ощущению вкуса. Когда пища или другие вещества попадают в рот, молекулы взаимодействуют со слюной и связываются со вкусовыми рецепторами в полости рта и других местах. Молекулы, придающие ощущение вкуса, считаются "разумными".
Вкусовые рецепторы делятся на две группы:
Визуальный, обонятельный, «сапиктивный» (восприятие вкусов), тройничный (горячий, прохладный), механический - все они способствуют восприятию вкуса. Из них транзиторные рецепторы потенциального катионного канала подсемейства V член 1 (TRPV1 ) ваниллоидные рецепторы отвечают за восприятие тепла некоторыми молекулами, такими как капсаицин, а рецептор CMR1 отвечает за восприятие холода такими молекулами, как ментол, эвкалиптол и ицилин.
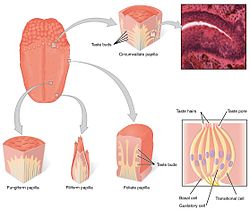
вкусовая система состоит из клеток вкусовых рецепторов в вкусовых рецепторах. Вкусовые рецепторы, в свою очередь, содержатся в структурах, называемых сосочками. Во вкусе участвуют три типа сосочков: грибовидные сосочки, листовые сосочки и кольцевые сосочки. (Четвертый тип - нитевидные сосочки вкусовых сосочков не содержат). Помимо сосочков, вкусовые рецепторы также находятся в нёбе и в ранних частях пищеварительной системы, таких как гортань и верхний пищевод. Есть три черепных нерва, которые иннервируют язык; блуждающий нерв, языкоглоточный нерв и лицевой нерв. языкоглоточный нерв и chorda tympani ветвь лицевого нерва иннервируют вкусовые рецепторы TAS1R и TAS2R. Наряду со вкусовыми рецепторами на языке эпителий кишечника также оснащен тонкой хемосенсорной системой, которая передает сенсорную информацию нескольким эффекторным системам, участвующим в регуляции аппетита, иммунных реакций и моторики желудочно-кишечного тракта
В В 2010 году исследователи обнаружили горькие рецепторы в легочной ткани, которые вызывают расслабление дыхательных путей при попадании горького вещества. Они считают, что этот механизм является эволюционно адаптивным, поскольку он помогает избавиться от инфекций легких, но также может быть использован для лечения астмы и хронической обструктивной болезни легких.
Вкус помогает идентифицировать токсины, поддерживают питание и регулируют аппетит, иммунные реакции и перистальтику желудочно-кишечного тракта. Сегодня различают пять основных вкусов: соленый, сладкий, горький, кислый и умами. Ощущения соленого и кислого вкуса обнаруживаются через ионные каналы. Сладкий, горький вкус и вкус умами, однако, обнаруживаются с помощью связанных с G-белком вкусовых рецепторов.
Кроме того, некоторые агенты могут действовать как модификаторы вкуса, как миракулин или куркулин для сладкого или стерубин на маскирующий горький.
Стандартный горький, сладкий, или вкусовой рецептор умами представляет собой G-белок-связанный рецептор с семью трансмембранными доменами. Связывание лиганда на вкусовых рецепторах активирует каскады второго мессенджера для деполяризации вкусовой клетки. Густдуцин - это наиболее распространенная вкусовая субъединица Gα, играющая главную роль в восприятии горького вкуса TAS2R. Густдуцин является гомологом трансдуцина, G-белка, участвующего в трансдукции зрения. Кроме того, вкусовые рецепторы используют ионный канал TRPM5, а также фосфолипазу PLCβ2.
TAS1R1 + Гетеродимерный рецептор TAS1R3 функционирует как рецептор умами, отвечая на связывание L- аминокислоты, особенно L- глутамата. Вкус умами чаще всего ассоциируется с пищевой добавкой глутамат натрия (MSG) и может быть усилен за счет связывания инозинмонофосфата (IMP) и молекулы гуанозинмонофосфата (GMP). Клетки, экспрессирующие TAS1R1 + 3, обнаруживаются в основном в грибовидных сосочках на кончике и краях языка и в клетках вкусовых рецепторов неба в нёбе. Показано, что эти клетки синапсируют с нервами барабанной перепонки, чтобы посылать свои сигналы в мозг, хотя была обнаружена некоторая активация языкоглоточного нерва.
Альтернативный кандидат Вкусовые рецепторы умами включают варианты сплайсинга метаботропных рецепторов глутамата, mGluR4 и mGluR1 и рецептор глутамат-ионного канала N-метил-D-аспартат типа <159.>
Гетеродимерный рецептор TAS1R2 + TAS1R3 функционирует как рецептор сладкого, связываясь с широкий ассортимент сахаров и заменителей сахара. Клетки, экспрессирующие TAS1R2 + 3, обнаружены в кольцевидных сосочках и листовых сосочках около задней части языка и нёба рецепторных клеток вкусовых рецепторов небо. Показано, что эти клетки действуют в синапс на chorda tympani и языкоглоточный нерв, чтобы посылать свои сигналы в мозг. Гомодимер TAS1R3 также действует как рецептор сладкого во многом так же, как TAS1R2 + 3, но имеет пониженную чувствительность к сладким веществам. Природные сахара легче обнаруживаются рецептором TAS1R3, чем заменителями сахара. Это может помочь объяснить, почему сахар и искусственные подсластители имеют разные вкусы. Генетический полиморфизм TAS1R3 отчасти объясняет разницу в восприятии сладкого вкуса и потреблении сахара между людьми афроамериканского происхождения и людьми европейского и азиатского происхождения.
Белки TAS2R (InterPro : IPR007960 ) функционируют как рецепторы горького вкуса. Существует 43 гена TAS2R человека, каждый из которых (за исключением пяти псевдогенов ) лишен интронов и кодирует белок GPCR. Эти белки, в отличие от белков TAS1R, имеют короткие внеклеточные домены и расположены в кольцевых сосочках, нёбе, листовых сосочках и надгортаннике <174.>вкусовые рецепторы с пониженной экспрессией в грибовидных сосочках. Хотя несомненно, что несколько TAS2R экспрессируются в одной клетке вкусового рецептора, все еще ведутся споры о том, могут ли млекопитающие различать вкусы разных горьких лигандов. Однако должно происходить некоторое перекрытие, поскольку горьких соединений гораздо больше, чем генов TAS2R. Обычные горькие лиганды включают циклогексимид, денатоний, PROP (6-н-пропил-2-тиоурацил ), PTC (фенилтиокарбамид ), и β- глюкопиранозиды.
Передача сигнала горьких стимулов осуществляется через α-субъединицу густдуцина. Эта субъединица G-белка активирует вкусовую фосфодиэстеразу и снижает уровни циклических нуклеотидов. Дальнейшие шаги на пути трансдукции пока неизвестны. Βγ-субъединица густдуцина также опосредует вкус, активируя IP 3(инозитолтрифосфат ) и DAG (диглицерид ). Эти вторичные мессенджеры могут открывать закрытые ионные каналы или вызывать высвобождение внутреннего кальция. Хотя все TAS2R расположены в клетках, содержащих густдуцин, нокаут густдуцина не устраняет полностью чувствительность к горьким соединениям, предлагая избыточный механизм горького вкуса (неудивительно, учитывая, что горький вкус обычно сигнализирует о присутствии токсина ). Один из предложенных механизмов независимого от густдуцина горького вкуса заключается в взаимодействии ионных каналов со специфическими горькими лигандами, аналогично взаимодействию ионных каналов, которое происходит при дегустации кислых и соленых стимулов.
Один из наиболее изученных белков TAS2R TAS2R38, что способствует дегустации как PROP, так и PTC. Это первый вкусовой рецептор, полиморфизм которого, как показано, ответственен за различия во вкусовом восприятии. Текущие исследования сосредоточены на определении других таких полиморфизмов, определяющих вкусовой фенотип. Более поздние исследования показывают, что генетический полиморфизм в других генах рецепторов горького вкуса влияет на восприятие горького вкуса кофеина, хинина и бензоата денатония.
Исторически считалось, что кислый вкус вызывается только тогда, когда свободные ионы водорода (H) непосредственно деполяризуют вкусовые рецепторы. Однако сейчас предлагаются специфические рецепторы кислого вкуса с другими методами действия. HCN1 и HCN4 (каналы HCN ) были двумя такими предложениями; оба этих рецептора представляют собой каналы, управляемые циклическими нуклеотидами. Предполагается, что два ионных канала вносят вклад в кислый вкус: ACCN1 и TASK-1.
Также были предложены различные рецепторы для соленого вкуса, наряду с возможным обнаружением вкуса липидов, сложных углеводов и воды. Однако доказательства наличия этих рецепторов в лучшем случае шатки и часто неубедительны в исследованиях на млекопитающих. Например, предложенный рецептор ENaC для обнаружения натрия может быть показан только как способствующий вкусу натрия при дрозофилии.
Фермент, связанный с кислым рецептором, передает информацию о газированной воде.
Идентифицирован возможный вкусовой рецептор жира, CD36. CD36 был локализован в окружных и листовидных сосочках, которые присутствуют в вкусовых сосочках и где продуцируется язычная липаза, и исследования показали, что рецептор CD36 связывает длинноцепочечные жирные кислоты. Различия в количестве экспрессии CD36 у людей были связаны с их способностью ощущать жиры, создавая аргументы в пользу связи рецептора с вкусом жира. Дальнейшие исследования рецептора CD36 могут быть полезны для определения существования настоящего рецептора вкуса жира. Предполагается, что
GPR120 и GPR40 реагируют на жир в полости рта, и их отсутствие приводит к снижению предпочтения жиров и снижению нейрональной реакции на пероральные жирные кислоты.
TRPM5 имеет было показано, что он участвует в реакции орального жира и идентифицирован как возможный рецептор орального жира, но недавние данные представляют его в первую очередь как нисходящий фактор.
Гены рецепторов горького вкуса человека названы TAS2R1 к TAS2R64, со многими пробелами из-за несуществующих генов, псевдогенов или предполагаемых генов, которые не были аннотированы в самой последней сборке генома человека. Многие гены рецепторов горького вкуса также имеют непонятные синонимы с несколькими разными названиями генов, относящимися к одному и тому же гену. См. Полный список генов рецепторов горького вкуса человека в таблице ниже:
| Класс | Ген | Синонимы | Псевдонимы | Локус | Описание |
|---|---|---|---|---|---|
| тип 1. (сладкий) | TAS1R1 | GPR70 | 1p36.23 | ||
| TAS1R2 | GPR71 | 1p36.23 | |||
| TAS1R3 | 1p36 | ||||
| тип 2. (горький) | TAS2R1 | 5p15 | |||
| TAS2R2 | 7p21.3 | псевдоген | |||
| TAS2R3 | 7q31. 3-q32 | ||||
| TAS2R4 | 7q31.3-q32 | ||||
| TAS2R5 | 7q31.3-q32 | ||||
| TAS2R6 | 7 | не аннотирован в сборке генома человека | |||
| TAS2R7 | 12p13 | ||||
| TAS2R8 | 12p13 | ||||
| TAS2R9 | 12p13 | ||||
| TAS2R10 | 12p13 | ||||
| TAS2R11 | отсутствует у людей | ||||
| TAS2R12 | TAS2R26 | 12p13.2 | псевдоген | ||
| TAS2R13 | 12p13 | ||||
| TAS2R14 | 12p13 | ||||
| TAS2R15 | 12p13.2 | псевдоген | |||
| TAS2R16 | 7q31.1-q31.3 | ||||
| TAS2R17 | отсутствует у человека | ||||
| TAS2R18 | 12p13.2 | псевдоген | |||
| TAS2R19 | TAS2R23, TAS2R48 | 12p13.2 | |||
| TAS2R20 | TAS2R49 | 12p13.2 | |||
| TAS2R21 | а отсутствует у человека | ||||
| TAS2R22 | 12 | не аннотирован в сборке генома человека | |||
| TAS2R24 | отсутствует у человека | ||||
| TAS2R25 | отсутствует у человека | ||||
| TAS2R27 | отсутствует у человека | ||||
| TAS2R28 | отсутствует у человека | ||||
| TAS2R29 | отсутствует у человека | ||||
| TAS2R30 | TAS2R47 | 12p13.2 | |||
| TAS2R31 | TAS2R44 | 12p13. 2 | |||
| TAS2R32 | отсутствует у человека | ||||
| TAS2R33 | 12 | не аннотирован в сборке генома человека | |||
| TAS2R34 | отсутствует у человека | ||||
| TAS2R35 | отсутствует в люди | ||||
| TAS2R36 | 12 | не аннотированы в сборке генома человека | |||
| TAS2R37 | 12 | не аннотированы в сборке генома человека | |||
| TAS2R38 | 7q34 | ||||
| TAS2R39 | 7q34 | ||||
| TAS2R40 | GPR60 | 7q34 | |||
| TAS2R41 | 7q34 | ||||
| TAS2R42 | 12p13 | ||||
| TAS2R43 | 12p13.2 | ||||
| TAS2R45 | GPR59 | 12 | |||
| TAS2R46 | 12p13.2 | ||||
| TAS2R50 | TAS2R51 | 12p13.2 | |||
| TAS2R52 | отсутствует у человека | ||||
| TAS2R53 | отсутствует у человека | ||||
| TAS2R54 | отсутствует у человека | ||||
| TAS2R55 | отсутствует у человека | ||||
| TAS2R56 | отсутствует у человека | ||||
| TAS2R57 | отсутствует у человека | ||||
| TAS2R58 | отсутствует у человека | ||||
| TAS2R59 | отсутствует у человека | ||||
| TAS2R60 | 7 | ||||
| TAS2R62P | 7q34 | псевдоген | |||
| TAS2R63P | 12p13.2 | псевдоген | |||
| TAS2R64P | 12p13.2 | псевдоген |
У многих видов вкусовые рецепторы показали потерю функций. Эволюционный процесс, в котором вкусовые рецепторы утратили свою функцию, считается адаптивной эволюцией, связанной с экологией кормления, определяющей специализацию и бифуркацию вкусовых рецепторов. Показано, что из всех вкусовых рецепторов горький, сладкий и умами имеют корреляцию между инактивацией вкусовых рецепторов и пищевым поведением. Однако нет убедительных доказательств того, что у каких-либо позвоночных отсутствуют гены рецепторов горького вкуса.
Рецептор сладкого вкуса является одним из вкусовых рецепторов, функция которого утрачена. У млекопитающих преобладающим рецептором сладкого вкуса является вкусовой рецептор типа 1 Tas1r2 / Tas1r3. Некоторые виды млекопитающих, такие как кошки и летучие мыши-вампиры, показали неспособность ощущать сладкий вкус. У этих видов причина потери функции сладкого рецептора связана с псевдогенизацией Tas1r2. Псевдогенизация Tas1r2 также наблюдается у видов, не относящихся к млекопитающим, таких как куры и безъязычная западная когтистая лягушка, и эти виды также демонстрируют неспособность ощущать сладкий вкус. Псевдогенизация Tas1r2 широко распространена и независима в отряде Carnivora. Многие исследования показали, что псевдогенизация вкусовых рецепторов вызвана вредной мутацией в открытых рамках считывания (ORF). В ходе исследования было обнаружено, что у хищных видов, отличных от кошачьих, эти виды демонстрировали мутации Tas1r2, нарушающие ORF, и они возникали независимо среди видов. Они также показали высокую вариативность в своих родословных. Предполагается, что псевдогенизация Tas1r2 произошла в результате конвергентной эволюции, когда хищные виды потеряли способность ощущать сладкий вкус из-за пищевого поведения.
Умами также является вкусовым рецептором, функция которого у многих видов потеряна. Преобладающими рецепторами вкуса умами являются Tas1r1 / Tas1r3. У двух линий водных млекопитающих, включая дельфинов и морских львов, было обнаружено, что Tas1r1 является псевдогенизированным. Псевдогенизация Tas1r1 также была обнаружена у наземных плотоядных видов. Хотя панда принадлежит к отряду Carnivora, она травоядна, где 99% ее рациона составляет бамбук, и она не может попробовать умами. Последовательность генома панды показывает, что ее ген Tas1r1 псевдогенизирован. В ходе исследования было обнаружено, что у всех видов отряда Carnivora, кроме панды, открытая рамка считывания сохранялась. У панды соотношение несинонимичных и синонимичных замен оказалось намного выше, чем у других видов отряда Carnivora. Эти данные коррелируют с датой окаменелостей панды, чтобы показать, где она перешла с диеты хищников на диету травоядных. Таким образом, предполагается, что потеря функции умами у панды вызвана изменением диеты, когда панда стала меньше зависеть от мяса. Однако эти исследования не объясняют травоядных животных, таких как лошади и коровы, которые сохранили рецептор Tas1r1.
В целом, потеря функции вкусового рецептора - это эволюционный процесс, который произошел из-за изменения рациона вида..