
Войти
| Tarsophlebiidae. Временной диапазон: 163–121 Ma PreꞒ Ꞓ O S D C P T J K Pg N Верхняя юра - Нижний мел | |
|---|---|
 | |
| Tarsophlebia eximia, верхняя юра, Solnhofen Plattenkalk | |
| Научная классификация | |
| Домен: | Eukaryota |
| Царство: | Animalia |
| Тип: | Arthropoda |
| Класс : | Insecta |
| Отряд: | Odonata |
| Подотряд: | †Tarsophlebioptera |
| Семейство: | †Tarsophlebiidae. Handlirsch, 1906 |
| Типовой род | |
| Tarsophlebia. Хаген, 1866 | |
| Род | |
| |
Tarsophlebiidae - это существующее семейство средних ископаемых одонат из верхней юры и нижнего мела периода Евразия. Они являются либо самым базальным представителем класса девица-стрекоза («анизозигоптеры») в пределах стебельной группы Anisoptera, либо сестринской группой всех современных стрекоз. Для них характерны базально открытая дискоидальная ячейка в обеих парах крыльев, очень длинные ноги, лопатообразные самцы cerci и гипертрофированный яйцеклад у самок.
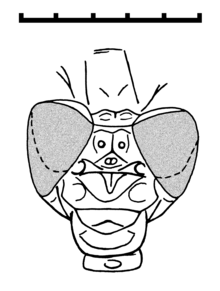 Tarsophlebia eximia, верхняя юра, Solnhofen Plattenkalk, голова самца экземпляра MCZ 6129
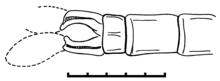
Tarsophlebia eximia, верхняя юра, Solnhofen Plattenkalk, голова самца экземпляра MCZ 6129  Tarsophlebia eximia, верхняя юра, Solnhofen Plattenkalk, правая задняя конечность голотипа
Tarsophlebia eximia, верхняя юра, Solnhofen Plattenkalk, правая задняя конечность голотипа Голова похожа на голову современных Gomphidae с двумя большими и шаровидными сложными глазами, которые четко разделены, но ближе друг к другу, чем у стрекоз (Zygoptera ). Есть также два головных шва.. Птероторакс, кажется, еще более перекошен, чем у стрекоз. Ноги очень длинные, с короткими сильными шипами и очень удлиненными лапками. Всего члеников предплюсны три (tarsomeres ), из которых самый базальный вдвое длиннее остальных. В паре коготков предплюсны отсутствует вентро-апикальный крючок современных стрекоз.
Самый крупный вид Turanophlebia sinica достиг размаха крыльев около 10,3 см (4,1 дюйма), в то время как самые маленькие виды Tarsophlebia minor достигли размаха крыльев только около 5,5 см (2,2 дюйма).
 Tarsophlebia minor, верхняя юра, Solnhofen Plattenkalk, заднее крыло голотипа, масштаб 10 мм
Tarsophlebia minor, верхняя юра, Solnhofen Plattenkalk, заднее крыло голотипа, масштаб 10 мм Жилкование крыла характеризуется следующими чертами:. крылья прозрачные, тонкие, без стеблей; дискоидная ячейка в основании обеих пар крыльев открыта, так что дуга неполная; дискоидная клетка переднего крыла очень острая; большая и острая субдисковидная ячейка заднего крыла; первичные антенные скобы Ax1 и Ax2 прочнее поперечных вторичных антенодальных вен; узелок в дистальном положении на 44-47% длины крыла; узелок с концевым перегибом ЦП и сильной узловой бороздой; птеростигма удлиненная (покрывающая несколько ячеек), с косой коровьей жилкой; одна лестиновая косая вена «О» присутствует между RP2 и IR2; во всех крыльях есть пары вторичных продольных вогнутых интеркалярных жилок, передних и задних выпуклых жилок CuA, MA и IR2, и близко им параллельных (задние интеркаляры всегда длиннее, чем связанные с ними передние интеркаляры); задние крылья без жилки CuAb; поперечно-жилковидный остаток жилки CuP изогнут и скорее похож на ветвь AA.
 Tarsophlebia eximia, верхняя юра, Solnhofen Plattenkalk, церки самца MCZ 6222
Tarsophlebia eximia, верхняя юра, Solnhofen Plattenkalk, церки самца MCZ 6222 Самцы различаются лопаточными церками, в то время как самки отличаются очень длинным и тонким гипертрофированным яйцекладом, который выступает далеко за пределы живота.
 Tarsophlebia eximia, верхняя юра, Solnhofen Plattenkalk, вторичный генитальный аппарат особи самца JME SOS 1720
Tarsophlebia eximia, верхняя юра, Solnhofen Plattenkalk, вторичный генитальный аппарат особи самца JME SOS 1720 Вторичные гениталии самцов принадлежали к уникальному примитивному типу, с маленьким семенным пузырьком на стерните 3, двумя парами небольших пластинчатых хамулей на стерните 2 и очень коротким срединным язычком на стерните 2. Очевидно, что ни одна из этих структур не гипертрофирована. интромитентный орган спермы (функциональный половой член). В каждом из трех подотрядов современных зубатых особей разные части этого аппарата увеличиваются и развиваются как интромитентный орган и устройство для удаления чужеродных сперматозоидов (конкуренция сперматозоидов): у Zygoptera это язычок, средний отросток стернита 2; у Epiophlebiidae - это латеральная пара задних ветвей на 2-м сегменте; а у Anisoptera это непарный семенной пузырек на передней части стернита 3. Строение аппарата Tarsophlebiidae наводит на мысль о промежуточном состоянии между протодонатами и современными стернитами. Исключительно хорошо сохранившийся самец Namurotypus sippeli показал, что у протодоната Meganisoptera полностью отсутствовал вторичный генитальный аппарат на 2-м и 3-м сегментах брюшка, а на 9-м сегменте все еще имелись первичные гениталии, которые сильно напоминают гениталии бескрылых чешуек, которые имеют не совокупляться, но откладывать внешние сперматофоры. Только на основе такого брачного поведения вообще возможна эволюция вторичного совокупления стрекоз. Первым шагом, вероятно, было прикрепление сперматофора к базальным стернитам брюшка самца вместо отложения на субстрате. Теперь самке нужно было извлечь сперматофор из вентральной полости самца. Это создало адаптивное давление, чтобы создать структуры хранения для сперматофора и структуры прикрепления для женских гениталий. Tarsophlebiidae, вероятно, представляют это состояние эволюции. Развитие жидкой спермы и интромитентных органов для совокупления, по-видимому, происходило три раза параллельно в трех существующих подотрядах.
Боковые ушные раковины по бокам базального брюшка у самцов Tarsophlebia eximia были описаны Nel et al. (1993). Однако Bechly (1996) показал, что эти предполагаемые мужские ушные раковины были основаны на неправильной интерпретации задних Hamuli, что было подтверждено Fleck et al. (2004).
Fleck et al. (2004) продемонстрировали, что самцы Tarsophlebiidae действительно обладают уникальным типом анальных придатков. Описание калоптеригоидных придатков Tarsophlebia eximia, по-видимому, с двумя парами кламмеров, было основано на неправильной интерпретации из-за артефактов сохранности. Церки очень длинные, с двустворчатым базальным черешком и дистальным пластинчатым расширением. Сломанные двухствольные черешки двух церок обычно ошибочно интерпретируются как две пары кламмеров, а дистальные пластинки игнорируются или рассматриваются как артефакты. Действительно, Tarsophlebia не имеет придатков, подобных зигоптероидам и анизоптероидам. Нет видимых парапроктов (как у Zygoptera) и нет эпипрокта (как у Epiophlebiidae и Anisoptera). Неясно, уменьшаются ли эти структуры вторично или отсутствуют в первую очередь.
Ископаемых личинок этого вымершего семейства еще не обнаружено.
Семейство было установлено Хандлиршем (1906) на основе типа рода Tarsophlebia Hagen, 1866.
Tarsophlebiopsis mayi был признан действительным большинством авторов до последнего пересмотра семейства Fleck et al. (2004), которые продемонстрировали, что голотип этого таксона, скорее всего, является всего лишь аберрантным экземпляром Tarsophlebia eximia.
Семейство содержит 2 описанных рода с 9 действительными видами:
Все остальные описанные таксоны являются синонимами типового вида Tarsophlebia eximia.
Монофилия Tarsophlebiidae убедительно подтверждается следующим набором производных признаков (аутапоморфии ):. задние крылья с гипертрофированной субдискоидальной клеткой, которая развивается как «псевдодискоидальная клетка»; слияние жилок MAb + MP + CuA на значительное расстояние до разделения MP и CuA в заднем крыле; жилка AA сильно изогнута в месте вставки CuP-пересечения; крайне острые дистальные углы дискоидальной и субдискоидальной ячеек переднего крыла. Признаки тела «отчетливо удлиненные ноги, с очень длинными лапками» и «мужские церки с лопаточными дистальными расширениями» известны от одного вида рода Tarsophlebia (T. eximia) и Turanophlebia (T. vitimensis) соответственно и, таким образом, принадлежали к общему наземному плану всех Tarsophlebiidae. Чрезвычайно удлиненный яйцеклад женского пола может быть еще одной синапоморфией для семьи, но он известен только по T. eximia и по единственному экземпляру T. minor в музее Зольнхофена.
Bechly (1996) и Nel et al. al. (2004: Приложение 2) предложил длинный список предполагаемых синапоморфий, которые демонстрируют более тесную связь Tarsophlebiidae с коронной группой Odonata:. наличие реберного треугольника в виде широкой и сильной склеротизации базального реберного края; дистальная дискоидальная вена MAb (= дистальная сторона дискоидальной ячейки) и субдискоидальная вена (происхождение CuA на MP) выровнены и дорсально усилены сильной склеротизацией, так что эта структура, кажется, пересекает вены MP и вогнутую складку вдоль эта вена (формирование «дискального корсета» sensu Carle 1982); эта дисковая скоба выровнена с аркулусом на плане (сохраняется только в некоторых таксонах с открытой дискоидальной ячейкой и в передних крыльях Epiophlebia); средняя вилка (первая вилка RP и основание IR2) смещена в основании, при этом RP3 / 4 обычно возникает из основания подузла (перевернутый у некоторых Coenagrionoidea), а RP2 возникает рядом с подузлом (на плане земли); более производный тип узлов, с перегибом в ScP; косая базальная скобка (все еще присутствует у Protanisoptera и Protozygoptera) трансформируется в поперечную «базальную скобку» Ax0, которая выглядит как первичная антенодальная поперечная жилка; наличие двух сильных первичных антенодальных поперечных жилок Ax1 и Ax2 (также присутствующих у некоторых Protozygoptera, например, экз. PIN 1/276 из верхней перми России); птеростигма отчетливо окаймлена косой постсубноузельной поперечной жилкой под базальным краем птеростигмы; наличие трахеированной лестиновой косой вены «О» между RP2 и IR2 (вторично отсутствует в некоторых таксонах); в срединном пространстве (= базальном пространстве) выпуклый остаток медиального стержня («рудиментарный CuA» sensu Fraser 1957) подавлен, поскольку он сливается с локтевым стержнем в общий медиокубитальный стержень (предполагаемое наличие этого след в голотипе Tarsophlebiopsis - это тератология или артефакт сохранения), сходный с некоторыми Protanisoptera, Triadophlebiomorpha и Protozygoptera.
Bechly (1996) предположил, что несколько уникальных симплезиоморфных особенностей всех Tarsophlebiidae указывают на то, что это семейство представляет сестринскую группу всех современных Odonata. Этими особенностями являются базально открытая дискоидальная ячейка в заднем крыле (вместо закрытой), что подразумевает неполную аркулус, (пока оспариваемое) наличие четырех тарсомеров равной длины (вместо трех) и очень примитивное состояние вторичных самцов. генитальный аппарат (например, ligula orimentary; vesicula spermalis, все еще очень короткий и плоский с очень широким пором) без какого-либо интромитентного органа. Поэтому Бечли рассмотрел сходство Tarsophlebiidae и Epiprocta, упомянутых Нелом и др. (1993), а именно: менее разделенные и относительно большие глаза, наличие двух головных швов и маленькие шипы на ногах (также присутствующие у Meganisoptera) как симплезиоморфии.
Однако, основываясь на кладистическом исследовании 14 знаков, Fleck et al. (2004) снова предположили, что Tarsophlebiidae может быть скорее сестринской группой клады Epiprocta, в которую входят Epiophlebiidae и Anisoptera. Тем не менее, этот результат имеет очень низкую статистическую поддержку и с тем же успехом может быть артефактом компьютерного алгоритма экономии, поскольку ни один из 14 символов не представляет однозначной синапоморфии для Tarsophlebiidae и Epiprocta.
Huang Nel (2009) представили убедительные доказательства из новой ископаемой останки Tarsophlebiidae из Китая, что число тарсомеров всего три, как у современных стрекоз, но что первый тарсомер примерно в два раза длиннее других. Bechly (1996) уже обсуждал эту возможность и предположил, что такой длинный базальный тарсомер, по сравнению с коротким базальным тарсомером у протодонатов и всех современных стрекоз, все еще может быть плезиоморфным состоянием, которое могло возникнуть в результате слияния двух базальных тарсомеров. Хуанг и Нель (2004) утверждают, что нет никаких доказательств такого слияния в окаменелостях и что удлинение первого сегмента также может быть производной особенностью (аутапоморфия) Tarsophlebiidae.
Две альтернативные филогенетические позиции Tarsophlebiidae
Гипотеза Fleck et al. (2004):
| Odonata |
| |||||||||||||||||||||||||||
Гипотеза Behlyn (1996)., 2007):
| Odonatoptera |
| |||||||||||||||||||||
Практически ничего не известно об экологии и поведении Tarsophlebiidae, но, вероятно, они были очень похожи на современных стрекоз. С их очень длинным яйцекладом, самки, вероятно, вставляли свои яйца в ил в воде или рядом с водой, как и современные Cordulegastridae. Примечательно, что две другие группы мезозойских зубатых (а именно Steleopteridae и) имели аналогично удлиненный яйцеклад..
Летопись окаменелостей этого семейства варьируется от верхней юры Казахстана и G в нижний мел Англии, Забайкалья, Монголии и Китая. Нет никаких записей о Tarsophlebiidae из каких-либо ископаемых мест за пределами Евразийского региона.
Первые окаменелости tarsophlebiid были образцами Tarsophlebia eximia из Верхней юры Solnhofen Plattenkalk в Германии, описанных как "Heterophlebia eximia" "и Euphaea longiventris Хагена (1862 г.). Более крупные версии этого семейства были предоставлены Nel et al. (1993) и Fleck et al. (2004).
Помимо важных типовых образцов, есть также несколько исключительно хорошо сохранившихся окаменелостей, которые вносят вклад в наши знания о морфологии Tarsophlebiidae. К ним относятся следующие особи:
| Wikimedia Commons имеет СМИ, относящиеся к Tarsophlebiidae. |