
Войти
 Споры, образующиеся в течение жизненного цикла спор.
Споры, образующиеся в течение жизненного цикла спор.  Свежий снег частично покрывает мох с грубыми стеблями (Brachythecium rutabulum ), произрастающий на прореженном гибридном черном тополе (Populus x canadensis ). Показана последняя стадия жизненного цикла мха , где спорофиты видны до распространения их спор: калиптра (1) все еще прикреплен к капсула (2). Также можно различить верхушки гаметофитов (3). На вставке показаны окружающие черные тополя, растущие на песчаных суглинках на берегу колка, с выделенной областью.
Свежий снег частично покрывает мох с грубыми стеблями (Brachythecium rutabulum ), произрастающий на прореженном гибридном черном тополе (Populus x canadensis ). Показана последняя стадия жизненного цикла мха , где спорофиты видны до распространения их спор: калиптра (1) все еще прикреплен к капсула (2). Также можно различить верхушки гаметофитов (3). На вставке показаны окружающие черные тополя, растущие на песчаных суглинках на берегу колка, с выделенной областью. В биологии, a спора - единица полового или бесполого размножения, которая может быть адаптирована для распространения и для выживания, часто в течение продолжительных периодов времени, в неблагоприятные условия. Споры составляют часть жизненных циклов многих растений, водорослей, грибов и простейших. Бактериальные споры не являются частью полового цикла, но представляют собой устойчивые структуры, используемые для выживания в неблагоприятных условиях. Споры Myxozoan выделяют амебулы в своих хозяевах для паразитарной инфекции, но также размножаются внутри хозяев посредством спаривания двух ядер внутри плазмодия, который развивается из амебулы.
Споры обычно гаплоид и одноклеточный и продуцируются мейозом в спорангии диплоид спорофита. При благоприятных условиях спора может развиться в новый организм, используя митотическое деление, производя многоклеточный гаметофит, который в конечном итоге продолжает производить гаметы. Две гаметы сливаются, образуя зиготу, которая развивается в новый спорофит. Этот цикл известен как чередование поколений.
Споры семенных растений производятся внутри, а мегаспоры (образующиеся внутри семяпочек) и микроспоры участвуют в образовании более сложных структур. которые образуют единицы распространения, семена и пыльцевые зерна.
Термин спора происходит от древнегреческого слова σπορά спора, означающего «семя, посев ", относящийся к σπόρος sporos," посев "и σπείρειν speirein," сеять ".
В просторечии разница между «спорой» и «гаметой » заключается в том, что спора прорастет и превратится в спорлинг, в то время как гамете необходимо объединиться с другой гаметой, чтобы сформировать зиготу перед дальнейшим развитием.
Основное различие между спорами и семенами как единицами распространения заключается в том, что споры одноклеточные, первая клетка гаметофита, тогда как семена содержат внутри себя развивающийся зародыш (многоклеточный спорофит следующего поколения), образованный слиянием мужской гаметы пыльцевой трубки с женской гаметой, образованной мегагаметофитом внутри семяпочки. Споры прорастают, давая начало гаплоидным гаметофитам, а семена прорастают, давая начало диплоидным спорофитам.
Сосудистые растения Споры всегда гаплоидны. Сосудистые растения являются либо гомоспористыми (или изоспористыми), либо гетероспористыми . Гомоспористые растения производят споры одного размера и типа.
Гетероспористые растения, такие как семенные растения, колосовидные, иголки и папоротники порядка Salviniales, производят споры двух разных размеров: большая спора (мегаспора), действующая как «женская» спора, и меньшая (микроспора), действующая как «мужская». Такие растения обычно дают начало двум видам спор внутри отдельных спорангиев: либо мегаспорангий, который продуцирует мегаспоры, либо микроспорангий, который продуцирует микроспоры. У цветковых растений эти спорангии встречаются в плодолистиках и пыльниках соответственно.
Грибы обычно образуют споры в результате полового или бесполого размножения. Споры обычно гаплоидны и превращаются в зрелых гаплоидных особей посредством митотического деления клеток (урединиоспоры и телиоспоры среди ржавчины являются дикариотическими). Дикариотические клетки возникают в результате слияния двух гаплоидных клеток гамет. Среди спорогенных дикариотических клеток кариогамия (слияние двух гаплоидных ядер) происходит с образованием диплоидной клетки. Диплоидные клетки подвергаются мейозу с образованием гаплоидных спор.
Споры можно классифицировать несколькими способами:
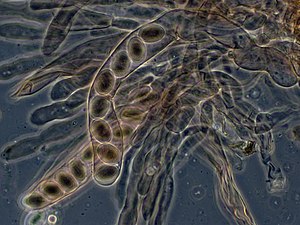 Asci из Morchella elata, содержащие аскоспоры
Asci из Morchella elata, содержащие аскоспоры  В растениях, микроспоры и в некоторых случаях мегаспоры образуются из всех четырех продуктов мейоза.
В растениях, микроспоры и в некоторых случаях мегаспоры образуются из всех четырех продуктов мейоза.  Напротив, во многих семенных растений и гетероспористых папоротников, только один продукт мейоза станет мегаспорой (макроспорой), а остальные дегенерируют.
Напротив, во многих семенных растений и гетероспористых папоротников, только один продукт мейоза станет мегаспорой (макроспорой), а остальные дегенерируют. У грибов и грибоподобных организмов споры часто классифицируются по структуре, в которой происходит мейоз и образование спор. Поскольку грибы часто классифицируют в соответствии с их спорообразующими структурами, эти споры часто характерны для определенного таксона из грибов.
Споры можно дифференцировать по тому, y может перемещать или нет.
 Ископаемые споры трилетов (синие) и тетрады спор (зеленые) позднесилурийского происхождения
Ископаемые споры трилетов (синие) и тетрады спор (зеленые) позднесилурийского происхождения  пыльца триколпатных растений Ricinus
пыльца триколпатных растений Ricinus При большом увеличении споры часто имеют сложные узоры или орнаменты на своей внешней поверхности. Для описания особенностей таких паттернов была разработана специальная терминология. Некоторые отметки представляют собой отверстия, места, где жесткая внешняя оболочка спор может проникнуть во время прорастания. Споры можно разделить на категории в зависимости от положения и количества этих отметин и отверстий. На спорах Alete нет линий. В спорах монолета на споре имеется единственная узкая линия (laesura). Это указывает на то, что материнская спора разделена на четыре части по вертикальной оси. В trilete spores каждая спора показывает три узкие линии, расходящиеся от центрального полюса. Это показывает, что четыре споры имели общее происхождение и первоначально находились в контакте друг с другом, образуя тетраэдр. Более широкое отверстие в форме канавки можно назвать colpus . По количеству колпий различают основные группы растений. Eudicots имеют трикольпатные споры (т.е. споры с тремя кольпи).
Тетрады спор, заключенные в оболочку, принимаются в качестве самые ранние свидетельства существования растений на суше относятся к середине ордовика (ранний Лланвирн, ~ 470 миллионов лет назад), то есть к периоду, из которого еще не были обнаружены макрофоссилии. Отдельные споры трилете, напоминающие споры современных криптогамных растений, впервые появились в летописи окаменелостей в конце ордовикского периода.
 Play media Споры, выбрасываемые грибами.
Play media Споры, выбрасываемые грибами. У грибов как бесполые, так и половые споры или спорангиоспоры многих видов грибов активно распространяются путем насильственного выброса из их репродуктивных структур. Этот выброс обеспечивает выход спор из репродуктивных структур, а также их перемещение по воздуху на большие расстояния. Таким образом, многие грибы обладают специализированными механическими и физиологическими механизмами, а также структурами на поверхности спор, такими как гидрофобины, для выброса спор. Эти механизмы включают, например, насильственный выброс аскоспор, обусловленный структурой аска, и накопление осмолитов в жидкостях аска, что приводит к взрывному выбросу аскоспор в воздух.
Насильственный выброс отдельных спор, называемых баллистоспорами, включает образование небольшой капли воды (капля Буллера ), которая при контакте со спорой приводит к ее высвобождению снаряда с начальным ускорением более 10 000 г. Другие грибы полагаются на альтернативные механизмы высвобождения спор, такие как внешние механические силы, примером которых являются клубочки. Привлечение насекомых, таких как мухи, к плодоносящим структурам, благодаря их яркому цвету и гнилостному запаху, для распространения спор грибов - еще одна стратегия, наиболее широко используемая вонючими рогами.
In Common Smoothcap moss (), вибрация спорофита, как было показано, является важным механизмом для высвобождения спор.
В случае распространения спор сосудистых растений, таких как папоротники, ветер Распределение очень легких спор обеспечивает большую способность к распространению. Кроме того, споры менее подвержены хищничеству животных, чем семена, потому что они почти не содержат запасов пищи; однако они более подвержены хищничеству грибков и бактерий. Их главное преимущество состоит в том, что из всех форм потомства для образования спор требуется меньше всего энергии и материалов.
В шиповнике Selaginella lepidophylla распространение частично достигается за счет необычного типа диаспоры, перекати-поле.

Споры мох Bartramia ithyphylla. (изображение под микроскопом, 400x)

Расщепленные спорангии папоротника. (вид под микроскопом, спор не видно)

Споры и элитеры хвоща. (Equisetum, вид под микроскопом)

Споры ископаемых растений (Scylaspora ) из силурийских отложений Швеции.

Плесень плода со спорами и заметным клеточным ростом. (2000x)

Скопления спор, сформированные внутри спорангиев слизистой плесени, из сосновых лесов востока Украины.
Внутренняя поверхность перидия слизистой плесени Tubifera dudkae со спорами.