
Войти
 Общие структуры сфинголипидов
Общие структуры сфинголипидов Сфингомиелин (SPH, ˌsfɪŋɡoˈmaɪəlɪn) является типом сфинголипидов, обнаруженных в мембранах клеток животных , особенно в мембранной миелиновой оболочке, которая окружает некоторые нервные клетки аксоны. Обычно он состоит из фосфохолина и церамида или головной группы фосфоэтаноламина ; следовательно, сфингомиелины также можно классифицировать как сфингофосфолипиды. У человека SPH составляет ~ 85% всех сфинголипидов и обычно составляет 10-20 мол.% Липидов плазматической мембраны.
Сфингомиелин был впервые выделен немецким химиком Иоганном Л.В. Thudicum в 1880-е гг. О структуре сфингомиелина впервые сообщили в 1927 году как о N-ацил-сфингозин-1-фосфорилхолине. Содержание сфингомиелина у млекопитающих колеблется от 2 до 15% в большинстве тканей, при этом более высокие концентрации обнаруживаются в нервных тканях, эритроцитах и линзах глаза. Сфингомиелин играет важную структурную и функциональную роль в клетке. Он является компонентом плазматической мембраны и участвует во многих сигнальных путях. Метаболизм сфингомиелина создает множество продуктов, которые играют важную роль в клетке.
 Сфингомиелин. Черный :Сфингозин. Красный :Фосфохолин. Синий: Жирная кислота
Сфингомиелин. Черный :Сфингозин. Красный :Фосфохолин. Синий: Жирная кислота  Вид сверху сфингомиелина, демонстрирующий его почти цилиндрическую форму
Вид сверху сфингомиелина, демонстрирующий его почти цилиндрическую форму Сфингомиелин состоит из головной группы фосфохолина, сфингозина и жирная кислота. Это один из немногих мембранных фосфолипидов, не синтезируемых из глицерина. Сфингозин и жирную кислоту вместе можно отнести к церамидам. Эта композиция позволяет сфингомиелину играть важную роль в сигнальных путях: деградация и синтез сфингомиелина производят важные вторичные мессенджеры для передачи сигнала.
Сфингомиелин, полученный из природных источников, таких как яйца или бычий мозг, содержит жирные кислоты с различной длиной цепи. Сфингомиелин с заданной длиной цепи, такой как пальмитоилсфингомиелин с насыщенной 16-ацильной цепью, коммерчески доступен.
В идеале молекулы сфингомиелина имеют форму цилиндра, однако многие молекулы сфингомиелина имеют значительное несоответствие цепей (длины двух гидрофобных цепей значительно различаются). Гидрофобные цепи сфингомиелина имеют тенденцию быть намного более насыщенными, чем другие фосфолипиды. Температура основной фазы перехода сфингомиелинов также выше по сравнению с температурой фазового перехода аналогичных фосфолипидов, около 37 ° С. Это может вносить латеральную неоднородность в мембрану, генерируя домены в бислое мембраны.
Сфингомиелин значительно взаимодействует с холестерином. Холестерин обладает способностью устранять фазовый переход из жидкого состояния в твердое в фосфолипидах. Поскольку температура перехода сфингомиелина находится в пределах физиологических температурных диапазонов, холестерин может играть важную роль в фазе сфингомиелина. Сфингомиелин также более склонен к межмолекулярным водородным связям, чем другие фосфолипиды.
Сфингомиелин синтезируется в эндоплазматическом ретикулуме (ER), где его можно найти в низких сумм и на транс-Гольджи. Он обогащен плазматической мембраной с большей концентрацией на внешнем, чем на внутреннем листке. Комплекс Гольджи представляет собой промежуточное звено между ER и плазматической мембраной, с немного более высокими концентрациями в направлении транс-стороны.
Синтез сфингомиелина включает ферментативный перенос фосфохолина от фосфатидилхолина до церамида. Первая обязательная стадия синтеза сфингомиелина включает конденсацию L-серина и пальмитоил-КоА. Эта реакция катализируется серинпальмитоилтрансферазой. Продукт этой реакции восстанавливается с образованием дигидросфингозина. Дигидросфингозин подвергается N-ацилированию с последующей десатурацией с образованием церамида. Каждая из этих реакций происходит на цитозольной поверхности эндоплазматического ретикулума. Церамид транспортируется в аппарат Гольджи, где он может быть преобразован в сфингомиелин. Сфингомиелинсинтаза отвечает за производство сфингомиелина из церамида. Диацилглицерин продуцируется как побочный продукт при переносе фосфохолина.
 Путь синтеза сфингомиелина de novo
Путь синтеза сфингомиелина de novo Распад сфингомиелина ответственен за запуск многих универсальных сигнальных путей. Он гидролизуется сфингомиелиназами (сфингомиелинспецифическими фосфолипазами типа C). Головная группа фосфохолина выделяется в водную среду, а церамид диффундирует через мембрану.
Мембранная миелиновая оболочка, которая окружает и электрически изолирует многие нервные клетки аксоны, особенно богата сфингомиелином, что предполагает его роль в качестве изолятора нервных волокон. плазматическая мембрана других клеток также богата сфингомиелином, хотя в основном он находится в экзоплазматическом листке клеточной мембраны. Однако есть некоторые свидетельства того, что во внутреннем листке мембраны также может находиться пул сфингомиелина. Более того, было обнаружено, что нейтральная сфингомиелиназа-2 - фермент, расщепляющий сфингомиелин на церамид - локализуется исключительно на внутренней створке, что также указывает на то, что там может присутствовать сфингомиелин.
Функция сфингомиелина оставалась неясной, пока не было обнаружено, что он играет роль в передаче сигнала. Было обнаружено, что сфингомиелин играет важную роль в сигнальных путях клеток. Синтез сфингомиелина на плазматической мембране сфингомиелинсинтазой 2 дает диацилглицерин, который представляет собой жирорастворимый вторичный мессенджер, который может проходить по сигнальному каскаду. Кроме того, распад сфингомиелина может производить церамид, который участвует в апоптотическом сигнальном пути.
Было обнаружено, что сфингомиелин играет роль в апоптозе клеток за счет гидролиза до церамида. Исследования, проведенные в конце 1990-х годов, показали, что церамид вырабатывается в различных условиях, ведущих к апоптозу. Затем была выдвинута гипотеза, что гидролиз сфингомиелина и передача сигналов церамидов важны для принятия решения о том, умирает ли клетка. В начале 2000-х годов появились новые исследования, которые определили новую роль гидролиза сфингомиелина в апоптозе, определяя не только, когда клетка умирает, но и как. После дополнительных экспериментов было показано, что если гидролиз сфингомиелина происходит на достаточно ранней стадии пути, производство церамида может влиять либо на скорость и форму гибели клеток, либо работать на высвобождение блоков при последующих событиях.
Сфингомиелин, а также другие сфинголипиды связаны с липидными микродоменами в плазматической мембране, известными как липидные рафты. Липидные рафты характеризуются тем, что липидные молекулы находятся в липидной упорядоченной фазе, предлагая большую структуру и жесткость по сравнению с остальной частью плазматической мембраны. В рафтах ацильные цепи имеют низкое движение цепи, но молекулы имеют высокую боковую подвижность. Этот порядок частично обусловлен более высокой температурой перехода сфинголипидов, а также взаимодействиями этих липидов с холестерином. Холестерин - это относительно небольшая неполярная молекула, которая может заполнять пространство между сфинголипидами, образованное большими ацильными цепями. Считается, что липидные рафты участвуют во многих клеточных процессах, таких как сортировка и транспортировка мембран, трансдукция сигналов и поляризация клеток. Избыток сфингомиелина в липидных рафтах может привести к инсулинорезистентности.
. Из-за специфических типов липидов в этих микродоменах липидные рафты могут накапливать определенные типы белков, связанных с ними, тем самым увеличивая особые функции, которыми они обладают. Предполагается, что липидные рафты участвуют в каскаде клеточного апоптоза.
Сфингомиелин может накапливаться при редком заболевании, называемом болезнью Ниманна – Пика, типы A и B. Это генетически наследуемое заболевание, вызванное дефицитом лизосомального фермента кислой сфингомиелиназы, которое вызывает накопление сфингомиелина в селезенке, печени, легких, костном мозге и мозге, вызывая необратимые неврологические повреждения. Из двух типов, включающих сфингомиелиназу, тип A встречается у младенцев. Он характеризуется желтухой, увеличенной печенью и глубоким повреждением мозга. Дети с этим типом редко доживают до 18 месяцев. Тип B включает увеличение печени и селезенки, что обычно происходит в подростковом возрасте. Мозг не поражен. У большинства пациентов <1% normal levels of the enzyme in comparison to normal levels. A hemolytic protein, lysenin, may be a valuable probe for sphingomyelin detection in cells of Niemann-Pick A patients.
в результате аутоиммунного заболевания рассеянного склероза (РС) миелиновая оболочка нейрональных клеток в головном и спинном мозге разрушается, что приводит к потере способности передавать сигнал. У пациентов с РС наблюдается повышенная регуляция некоторых цитокинов в спинномозговой жидкости, в частности, фактора некроза опухоли альфа. Это активирует сфингомиелиназу, фермент, катализирующий гидролиз сфингомиелина до церамида; Активность сфингомиелиназы наблюдалась в сочетании с клеточным апоптозом.
Избыток сфингомиелина в мембране эритроцитов (как в абеталипопротеинемии ) вызывает избыточное накопление липидов во внешней створке эритроцит плазматическая мембрана. В результате образуются эритроциты неправильной формы, называемые акантоцитами.
Шаровидная модель сфингомиелина
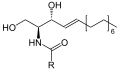
Скелетная формула сфингомиелина
.