
Войти
| Ооцит | |
|---|---|
| Идентификаторы | |
| MeSH | D009865 |
| FMA | 18644 |
| Анатомическая терминология [редактировать в Викиданных ] | |
An ооцит (UK :, US : ), ооцит, овоцит или редко ооцит, является самкой гаметоцит или половая клетка, участвующая в воспроизводстве. Другими словами, это незрелая яйцеклетка или яйцеклетка. Ооцит продуцируется в яичнике во время женского гаметогенеза. Женские половые клетки продуцируют первичные зародышевые клетки (PGC), которые затем подвергаются митозу, образуя оогонии. Во время оогенеза оогонии становятся первичными ооцитами. Ооцит - это форма генетического материала, который можно собирать для криоконсервации. Криоконсервация генетических ресурсов животных была введена в действие как средство сохранения традиционного домашнего скота.
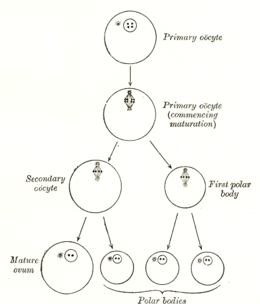 Диаграмма, показывающая уменьшение количества хромосом в процессе созревания яйцеклетки ; этот процесс известен как мейоз.
Диаграмма, показывающая уменьшение количества хромосом в процессе созревания яйцеклетки ; этот процесс известен как мейоз.Образование ооцита называется ооцитогенезом, который является частью оогенеза. Оогенез приводит к образованию как первичных ооцитов во время плода, так и вторичных ооцитов после него как часть овуляции.
| Тип клеток | плоидность / хромосомы | хроматиды | Процесс | Время завершения |
| Oogonium | диплоид / 46 (2N) | 2C | Ооцитогенез (митоз ) | третий триместр |
| первичный ооцит | диплоид / 46 (2N) | 4C | Оотидогенез (мейоз I) (Фолликулогенез ) | Диктиат в профазе I до 50 лет |
| вторичный ооцит | гаплоид / 23 (1N) | 2C | оотидогенез (мейоз II) | остановлен в метафазе II до оплодотворения |
| яйцеклетки | гаплоид / 23 (1N) | 1C | Оотидогенез (мейоз II) | Минуты после оплодотворения |
| Яйцо | гаплоид / 23 (1N) | 1C |
Ооциты богаты цитоплазмой, которая содержит гранулы желтка для питания клетки на ранней стадии развития.
Во время первичная стадия ооцитов оогенеза, th Ядро называется зародышевым пузырьком.
Единственный нормальный человеческий тип вторичного ооцита имеет 23-ю (половую) хромосому как 23, X (определяющий самку), тогда как сперматозоид может иметь 23, X (определяющий самку).) или 23, Y (определяется мужчиной).
Пространство внутри яйцеклетки или незрелой яйцеклетки находится в ячейке-гнезде.
Комплекс кумулюс-ооцит содержит слои плотно упакованных кумулюсных клеток, окружающих ооцит в фолликуле Граафа. Ооцит задерживается в Meiosis II на стадии метафазы II и считается вторичным ооцитом. Перед овуляцией кучевой комплекс претерпевает структурные изменения, известные как расширение кучевых облаков. Клетки гранулезы трансформируются из плотно уплотненного в расширенный слизистый матрикс. Многие исследования показывают, что расширение кумулюсов имеет решающее значение для созревания ооцита, поскольку комплекс кумулюсов является прямым связующим звеном ооцита с окружающей средой развивающегося фолликула. Он также играет важную роль в оплодотворении, хотя механизмы не совсем известны и зависят от вида.
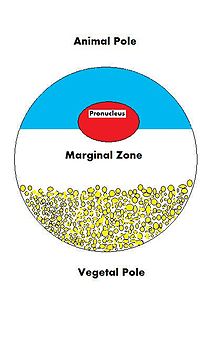 Полюса ооцитов
Полюса ооцитов Потому что судьба ооцита - оплодотворяться и в конечном итоге расти в полностью функционирующий организм, он должен быть готов регулировать множество клеточных процессов и процессов развития. Ооцит, большая и сложная клетка, должен быть снабжен многочисленными молекулами, которые будут направлять рост эмбриона и контролировать клеточную активность. Поскольку ооцит является продуктом женского гаметогенеза, вклад матери в ооцит и, следовательно, в вновь оплодотворенную яйцеклетку огромен. Есть много типов молекул, которые поставляются в ооцит из материнской цепи, которые будут направлять различные активности в растущей зиготе.
ДНК клетки уязвима на повреждающее действие окислительных свободных радикалов, образующихся как побочные продукты клеточного метаболизма. Повреждение ДНК, происходящее в ооцитах, если его не исправить, может привести к летальному исходу и привести к снижению плодовитости и потере потенциального потомства. Ооциты значительно больше средней соматической клетки, поэтому для их обеспечения необходима значительная метаболическая активность. Если бы эта метаболическая активность осуществлялась собственным метаболическим механизмом ооцита, геном ооцита подвергался бы действию образовавшихся реактивных побочных продуктов окисления. Таким образом, похоже, что процесс развился, чтобы избежать этой уязвимости ДНК зародышевой линии. Было высказано предположение, что во избежание повреждения ДНК генома ооцитов метаболизм, способствующий синтезу большей части компонентов ооцита, был перенесен на другие материнские клетки, которые затем перенесли эти компоненты в ооциты. Таким образом, ооциты многих организмов защищены от окислительного повреждения ДНК, в то же время накапливая большую массу веществ, питающих зиготу в ее первоначальном эмбриональном росте.
Во время роста ооцита различные материнские транскрибируемые информационные РНК или мРНК поставляются материнскими клетками. Эти мРНК могут храниться в комплексах мРНП (рибонуклеопротеин сообщения) и транслироваться в определенные моменты времени, они могут быть локализованы в определенной области цитоплазмы или могут быть гомогенно диспергированы в цитоплазме всего ооцита. Материнские загруженные белки также могут быть локализованы или повсеместно распространены по всей цитоплазме. Транслируемые продукты мРНК и загруженные белки выполняют множество функций; от регулирования клеточного «домашнего хозяйства», такого как развитие клеточного цикла и клеточный метаболизм, до регулирования процессов развития, таких как оплодотворение, активация зиготической транскрипции и формирование осей тела. Ниже приведены некоторые примеры унаследованных от матери мРНК и белков, обнаруженных в ооцитах африканской когтистой лягушки.
| Имя | Тип материнской молекулы | Локализация | Функция |
|---|---|---|---|
| VegT | мРНК | Растительное полушарие | Фактор транскрипции |
| Vg1 | мРНК | Растительное полушарие | Фактор транскрипции |
| XXBP-1 | мРНК | Неизвестно | Фактор транскрипции |
| CREB | Белок | Повсеместно | Фактор транскрипции |
| FoxH1 | мРНК | Повсеместно | Фактор транскрипции |
| p53 | Белок | Повсеместно | Фактор транскрипции |
| Lef/Tcf | мРНК | Повсеместно | Фактор транскрипции |
| FGF2 | Белок | Ядро | Неизвестно |
| FGF2, 4, 9 FGFR1 | мРНК | Неизвестно | Передача сигналов FGF |
| Эктодермин | Белок | Полушарие животных | Убиквитинлигаза |
| PACE4 | мРНК | Растительный полушарие | пропротеинконвертаза |
| Coco | Белок | Неизвестно | Ингибитор BMP |
| витая гаструляция | Белок | Неизвестно | BMP / Chordin-связывающий белок |
| fatvg | мРНК | Растительное полушарие | Формирование зародышевых клеток и вращение коры |
 Материнские детерминанты в Xenopus laevis ооците
Материнские детерминанты в Xenopus laevis ооците Ооцит получает митохондрии от материнских клеток, которые в дальнейшем будут контролировать эмбриональный метаболизм и апоптотические события. Разделение митохондрий осуществляется системой микротрубочек, которые будут локализовать митохондрии по всему ооциту. У некоторых организмов, таких как млекопитающие, отцовские митохондрии, доставленные сперматозоидом в ооцит, разрушаются за счет присоединения убиквитинированных белков. Разрушение отцовских митохондрий обеспечивает строго материнское наследование митохондрий и митохондриальной ДНК или мтДНК.
У млекопитающих ядрышко ооцита происходит исключительно от материнского клетки. Ядрышко, структура, обнаруженная внутри ядра, является местом, где рРНК транскрибируется и собирается в рибосомы. Хотя ядрышко плотное и неактивное в зрелом ооците, оно необходимо для правильного развития эмбриона.
Материнские клетки также синтезируют и вносят запас рибосом которые необходимы для трансляции белков до активации зиготического генома. В ооцитах млекопитающих рибосомы материнского происхождения и некоторые мРНК хранятся в структуре, называемой цитоплазматической решеткой. Эти цитоплазматические решетки, сеть из фибрилл, белка и РНК, увеличиваются в плотности по мере уменьшения количества рибосом в растущем ооците.
сперматозоид, оплодотворяющий ооцит, внесет свой пронуклеус, другую половину зиготического генома. У некоторых видов сперматозоиды также вносят вклад в центриоль, которая помогает образовывать зиготическую центросому, необходимую для первого деления. Однако у некоторых видов, например у мышей, вся центросома приобретается по материнской линии. В настоящее время изучается возможность других цитоплазматических вкладов сперматозоидов в эмбрион.
Во время оплодотворения сперма обеспечивает три основных части ооцита: (1) сигнальный или активирующий фактор, который вызывает активацию метаболически спящего ооцита; (2) гаплоидный отцовский геном; (3) центросома, которая отвечает за поддержание системы микротрубочек. См. Анатомию сперматозоидов
| Предшествующий. Нет | Этапы человеческое развитие. Сперма + ооцит | Преемник. зигота |
{kind=link}