
Войти
| Schistosoma | |
|---|---|
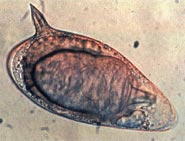 | |
| Яйцо Schistosoma mansoni | |
| Научная классификация | |
| Царство: | Animalia |
| Тип: | Platyhelminthes |
| Класс: | Trematoda |
| Отряд: | Diplostomida |
| Семейство: | Schistosomatidae |
| Род: | Schistosoma . Weinland, 1858 |
| Виды | |
Schistosoma bomfordi. Schistosoma bovis. Schistosoma curassoni. Schistosoma datta. Schistosoma edwardiense. Schistosoma guineensis. Schistosoma haematobium. Schistosoma402inasutis indicum. Schistosoma intercalatum. Schistosoma japonicum. Schistosoma kisumuensis. Schistosoma leiperi. Schistosoma malayensis. Schistosoma mansoni. Schistosoma margrebowiei. Schistosoma margrebowiei. Schistosoma434 ovuncatum. Schistosoma nasale. Schistosoma rodhaini. Schistosoma sinensium. Schistosoma spindale. Schistosoma turkestanicum | |
Schistosoma - это род из трематод, широко известных как кровяные сосальщики . Это паразитарные плоские черви, ответственные за очень значительную группу инфекций у людей под названием шистосомоз, которая считается по оценке Всемирной организации здравоохранения как второе по размеру социально-экономическое паразитарное заболевание (после малярии ), с сотнями миллионов инфицированных во всем мире.
Взрослые плоские черви паразитируют в кровеносных капиллярах брыжейки или сплетения мочевого пузыря, в зависимости от вида заражения. Они уникальны среди трематод и любых других плоских червей тем, что они раздельнополые с отчетливым половым диморфизмом между самцом и самкой. Тысячи яиц высвобождаются и достигают либо мочевого пузыря, либо кишечника (в зависимости от вида заражения), а затем выводятся с мочой или фекалиями в пресную воду. Личинки должны затем пройти через промежуточную улитку хозяин, прежде чем появится следующая личиночная стадия паразита, которая сможет заразить нового хозяина-млекопитающего, непосредственно проникая через кожу.
 Электронная микрофотография взрослого самца паразитирующего червя Schistosoma. Полоса (слева внизу) представляет длину 500 мкм.
Электронная микрофотография взрослого самца паразитирующего червя Schistosoma. Полоса (слева внизу) представляет длину 500 мкм. Происхождение этого рода остается неясным. В течение многих лет считалось, что этот род имеет африканское происхождение, но секвенирование ДНК предполагает, что виды (S. edwardiense и S. hippopotami), заражающие бегемота (Hippopotamus amphibius ) мог быть базальным. Поскольку гиппопотамы существовали как в Африке, так и в Азии в кайнозойскую эру, род, возможно, возник как паразиты бегемотов. Первоначальными хозяевами для видов Юго-Восточной Азии, вероятно, были грызуны.
. Основываясь на филогенетике улиток-хозяев, кажется вероятным, что этот род развился в Гондване между 70 миллионов лет назад и 120 миллионов лет назад.
сестринская группа Schistosoma - это род слонов, заражающих шистосомы - Bivitellobilharzia.
Паразиты крупного рогатого скота, овец, коз и кашемировых коз связаны с африканскими шистосомы. Этот последний вид с тех пор был переведен в род Schistosoma.
Внутри группы haematobium S. bovis и S. curassoni, по-видимому, тесно связаны между собой, как и S. leiperi и S. mattheei.
С. mansoni, по-видимому, возникла в Восточной Африке 0,43–0,30 миллиона лет назад.
S. mansoni и S. rodhaini, по-видимому, имели общего предка 107,5–147,6 тыс. лет назад. Этот период совпадает с самыми ранними археологическими свидетельствами рыболовства в Африке. Похоже, что S. mansoni возник в Восточной Африке, а эффективная численность популяции сократилась 20-90 тысяч лет назад, прежде чем расселиться по континенту в голоцене. Позже этот вид был передан в Америку в результате работорговли.
С. incognitum и S. nasale более тесно связаны с африканскими видами, чем с группой japonicum.
S. sinensium, по всей видимости, излучался в течение плиоцена.
S. mekongi, по-видимому, вторгся в Юго-Восточную Азию в середине плейстоцена.
Предполагаемые даты видообразования для группы japonicum : ~ 3,8 миллиона лет назад для S. japonicum / Шистосома Юго-Восточной Азии и ~ 2,5 миллиона лет назад для S. malayensis / S. mekongi.
Schistosoma turkestanicum заражает благородных оленей в Венгрии. Эти штаммы, по-видимому, отличаются от штаммов, обнаруженных в Китае и Иране. Дата расхождения, по-видимому, произошла за 270 000 лет до настоящего времени.
Род Schistosoma, как в настоящее время определяется, является парафилетическим, поэтому возможны пересмотры. В этом роде признано более двадцати видов.
Род был разделен на четыре группы: indicum, japonicum, haematobium и mansoni . Родство остальных видов все еще выясняется.
Тринадцать видов обитают в Африке. Двенадцать из них разделены на две группы - с боковым шипом на яйце (группа mansoni ) и с конечным шипом (группа haematobium ).
Четыре вида группы mansoni : S. edwardiense, S. hippotami, С. mansoni и С. rodhaini.
Девять видов группы haematobium: S. bovis, С. curassoni, С. guineensis, S. haematobium, S. intercalatum, S. kisumuensis, S. leiperi, С. margrebowiei и С. mattheei.
С. leiperi и S. matthei, по-видимому, связаны. S. margrebowiei базальна в этой группе. S. guineensis является родственным видом группировок S. bovis и S. curassoni. S. intercalatum на самом деле может быть комплексом видов по крайней мере из двух видов.
группа индика включает три вида: S. indicum, S. nasale и S. Спиндейл. Эта группа, по-видимому, появилась в плейстоцене. Все используют легочных улиток в качестве хозяев. S. spindale широко распространен в Азии, но встречается также в Африке. Они встречаются в Азии и Индии.
ю. indicum встречается в Индии и Таиланде..
Группа indicum, по-видимому, является сестринской кладой африканских видов.
Группа japonicum включает пять видов: S. japonicum, S. malayensis и S. меконги, С. ovuncatum и S. sinensium, и эти виды встречаются в Китае и Юго-Восточной Азии.
S. ovuncatum образует кладу с S. sinensium и встречается в северном Таиланде. Окончательный хозяин неизвестен, а промежуточный хозяин - улитка Tricula bollingi. Известно, что этот вид использует в качестве хозяев улиток семейства Pomatiopsidae.
S. incognitum, по-видимому, является базальным в этом роде. Это может быть более тесно связано с афро-индийскими видами, чем с группой Юго-Восточной Азии. Этот вид использует в качестве хозяев легочных улиток. Исследование митохондрий позволяет предположить, что Schistosoma incognitum может быть комплексом видов.
По состоянию на 2012 г. в этот род были переданы еще четыре вида, ранее классифицированных как виды в этом роде. Orientobilharzia. Orientobilharzia морфологически отличается от Schistosoma только количеством семенников. Обзор морфологических и молекулярных данных показал, что различия между этими родами слишком малы, чтобы оправдать их разделение. Это четыре вида:
Гибрид S. haematobium-S.guineenis наблюдалась в Камеруне в 1996 году. S. haematobium смог прижиться только после обезлесения тропических лесов в Лум рядом с эндемичным S. guineensis; гибридизация привела к конкурентному исключению S. guineensis.
В 2003 г. S. mansoni-S. Родхаини гибрид был обнаружен у улиток в западной Кении. По состоянию на 2009 год он не был обнаружен у людей.
В 2009 году S. haematobium – S. bovis были описаны у детей северного Сенегала. Бассейн реки Сенегал сильно изменился с 1980-х годов после строительства плотины Диама в Сенегале и плотины Манантали в Мали. Плотина Диама препятствовала проникновению океанской воды и открывала новые формы ведения сельского хозяйства. Миграция людей, увеличение количества домашнего скота и мест, где люди и крупный рогатый скот загрязняют воду, способствовали смешиванию различных шистосом в Nder, например. Этот же гибрид был идентифицирован в 2015 году в ходе расследования вспышки шистосомоза на Корсике, относящейся к реке Cavu.
В 2019 году S. haematobium – S. Гибрид mansoni был описан у 14-летнего пациента с гематурией из Кот-д'Ивуара.
A кладограмма на основе 18S рибосомной РНК, 28S рибосомная РНК и частичные гены субъединицы I цитохром с оксидазы (COI) показывают филогенные отношения видов в роде Schistosoma:
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


Географические области, связанные с шистосомозом Всемирной организацией здравоохранения по состоянию на январь 2017 года, включают в алфавитном порядке: Африку, Бразилию, Камбоджу, Карибский бассейн, Китай, Корсику, Индонезия, Лаос, Ближний Восток, Филиппины, Суринам и Венесуэла. В Европе не было случаев заболевания с 1965 года, пока не произошла вспышка на Корсике.
Паразитические плоские черви Schistosoma вызывают группу хронических инфекций, называемых шистосомозом. также как бильгарциоз. Препарат против шистосомы - это шистосомицид.
Паразитизм людей, вызываемый шистосомой, по-видимому, развивался как минимум трижды в Азии и Африке..
| Научное название | Первый промежуточный хозяин | Эндемическая зона |
|---|---|---|
| Schistosoma guineensis | Bulinus forskalii | Западная Африка |
| Schistosoma intercalatum | Bulinus spp | Африка |
| Schistosoma haematobium | Bulinus spp. | Африка, Ближний Восток |
| Schistosoma japonicum | Oncomelania spp. | Китай, Восточная Азия, Филиппины |
| Schistosoma malayensis | Robertsiella spp. | Юго-Восточная Азия |
| Schistosoma mansoni | Biomphalaria spp. | Африка, Южная Америка, Карибский бассейн, Ближний Восток |
| Schistosoma mekongi | Neotricula aperta | Юго-Восточная Азия |
Schistosoma indicum, Schistosoma nasale, Schistosoma spindale, Schistosoma leiperi являются паразитами жвачных.
Schistosoma edwardiense и являются паразитами бегемота.
Schistosoma ovuncatum и Schistosoma sinensium паразиты грызунов.
Взрослые шистосомы разделяют все основные черты дигенеи. У них основная двусторонняя симметрия, ротовые и вентральные присоски, тело покрыто синцитиальным тегументом, пищеварительной системой со слепым концом состоящий из рта, пищевода и раздвоенной слепой кишки ; область между тегументом и пищеварительным каналом заполнена рыхлой сеткой из мезодермы клеток и выделительной или осморегуляторной системой, основанной на клетках пламени. Взрослые черви обычно имеют длину 10–20 мм (0,39–0,79 дюйма) и используют глобины из гемоглобина своих хозяев для собственной системы кровообращения.
 В отличие от большинства плоских червей, шистосомы гонохористы. Можно увидеть узкую самку, выходящую из более толстого гинекофорального канала самца ниже его вентральной присоски.
В отличие от большинства плоских червей, шистосомы гонохористы. Можно увидеть узкую самку, выходящую из более толстого гинекофорального канала самца ниже его вентральной присоски. В отличие от других трематод, шистосомы раздельнополые, т. Е. Полы раздельны. Оба пола демонстрируют сильную степень полового диморфизма, и самец значительно крупнее самки. Самец окружает самку и заключает ее в свой гинакофорный канал на всю взрослую жизнь червей. Когда самец питается кровью хозяина, он передает часть ее самке. Самец также передает химические вещества, которые завершают развитие самки, после чего они размножаются половым путем. Хотя редко, но иногда спаривающиеся шистосомы «разводятся», при этом самка оставляет самца для другого самца. Точная причина не выяснена, хотя считается, что самки оставят своих партнеров, чтобы спариваться с более генетически далекими самцами. Такой биологический механизм мог бы служить для уменьшения инбридинга и может быть фактором необычно высокого генетического разнообразия шистосом.
Геномы Schistosoma haematobium, S. japonicum и С. mansoni.
Яйца этих паразитов впервые были замечены Теодором Максимилианом Бильгарцем, Немецкий патолог, работавший в Египте в 1851 году, обнаруживший яйца Schistosoma haematobium в ходе вскрытия. Он написал два письма своему бывшему учителю фон Зибольду в мае и августе 1851 года, в которых описал свои открытия. Фон Зибольд опубликовал статью в 1852 году, в которой резюмировал открытия Бильгарца и назвал червей Distoma haematobium. Бильхарц написал статью в 1856 году, в которой более полно описал червей. Их необычная морфология означала, что их нельзя было с комфортом включить в Дистому. Поэтому в 1856 году Меккель фон Хельмсбак (de ) создал для них род Bilharzia. В 1858 г. Давид Фридрих Вайнланд предложил название Schistosoma (греч. «Расщепленное тело»), потому что черви не были гермафродитами, а имели разные полы. Несмотря на то, что Bilharzia имеет приоритет, род название Schistosoma был официально принят Международной комиссией по зоологической номенклатуре. Термин Bilharzia для описания заражения этими паразитами до сих пор используется в медицинских кругах.
Бильгарц также описал Schistosoma mansoni, но этот вид был переописан Луи Вестенра Самбон в 1907 году в Лондонской школе тропической медицины, назвавшей его в честь своего учителя Патрика Мэнсона.
. В 1898 году все известные тогда виды были помещены в подсемейство Стайлзом и Хассель. В 1899 году Лосс повысил это до статуса семьи. В 1907 году Поче исправил грамматическую ошибку в фамилии. Жизненный цикл был определен бразильским паразитологом Пираджа да Силва (1873-1961) в 1908 году.
В 2009 году геномы Schistosoma mansoni и Расшифровка Schistosoma japonicum открыла путь для новых целевых методов лечения. В частности, исследование обнаружило, что геном S. mansoni содержит 11 809 генов, в том числе многие из них вырабатывают ферменты для расщепления белков, позволяя паразиту размножаться через ткань. Кроме того, S. mansoni не имеет фермента для производства определенных жиров, поэтому для их производства он должен полагаться на своего хозяина.
| Викискладе есть медиафайлы, связанные с Schistosoma. |