
Войти
| рибонуклеозид-дифосфатредуктаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Гетерооктамер рибонуклеозид-дифосфатредуктазы, E.Coli Гетерооктамер рибонуклеозид-дифосфатредуктазы, E.Coli | |||||||||
| Идентификаторы | |||||||||
| ЕС нет. | 1.17.4.1 | ||||||||
| № CAS | 9047-64-7sy | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| BRENDA | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| Структуры PDB | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | Amigo / QuickGO | ||||||||
| |||||||||
Рибонуклеотидредуктаза ( RNR), также известная как рибонуклеозиддифосфатредуктаза ( rNDP), представляет собой фермент, который катализирует образование дезоксирибонуклеотидов из рибонуклеотидов. Он катализирует это образование, удаляя 2'-гидроксильную группу рибозного кольца нуклеозиддифосфатов. Это восстановление производит дезоксирибонуклеотиды. Дезоксирибонуклеотиды, в свою очередь, используются в синтезе ДНК. Реакция, катализируемая RNR, строго сохраняется у всех живых организмов. Кроме того, RNR играет критическую роль в регулировании общей скорости синтеза ДНК, так что соотношение ДНК и массы клетки поддерживается на постоянном уровне во время деления клеток и репарации ДНК. Несколько необычная особенность фермента RNR заключается в том, что он катализирует реакцию, протекающую по свободнорадикальному механизму действия. Субстратами для RNR являются ADP, GDP, CDP и UDP. dTDP (дезокситимидиндифосфат) синтезируется другим ферментом ( тимидилаткиназой ) из dTMP (дезокситимидинмонофосфат).
Рибонуклеотидредуктазы делятся на три класса. Ферменты RNR класса I состоят из большой альфа-субъединицы и малых бета-субъединиц, которые связываются с образованием активного гетеродимерного тетрамера. Уменьшая NDP до 2'-dNDP, фермент катализирует синтез de novo дезоксирибонуклеотидов (dNTP), которые являются предшественниками синтеза ДНК и необходимы для пролиферации клеток. RNR класса II образуют 5'-дезоксиаденозильный радикал путем гомолитического расщепления связи C-Co в аденозилкобаламине. Кроме того, RNR класса III содержат стабильный глицильный радикал.
Люди несут RNR класса I. Альфа-субъединица кодируется геном RRM1, в то время как существуют две изоформы бета-субъединицы, кодируемые генами RRM2 и RRM2B:
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Каждый альфа- мономер I класса состоит из трех доменов :
В Pfam второй домен интерпретируется как два отдельных домена:
|
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Субъединица бета класса I обычно содержит биметаллический центр и стабильный тирозильный радикал. У человека бета-субъединица зависит от кофактора ди-железа. В E. coli тирозильный радикал расположен в положении 122 (Y122), обеспечивая стабильный радикал для субъединиц RNR2 класса I. У A. aegypti этот тирозильный радикал находится в положении 184 (Y184). Тирозильный радикал глубоко погружен внутри белка в гидрофобную среду, расположенную близко к центру железа, который используется для стабилизации тирозильного радикала. В структуре двух μ-оксо-связанных железов преобладают лиганды, которые служат сайтами связывания железа: четыре карбоксилата [ аспартат (D146), глутамат (E177, E240 и E274)] и два гистидина (H180 и H277). Ассоциация происходит между C-концом RNR2 и C-концом RNR1. Ферментативная активность зависит от ассоциации субъединиц RNR1 и RNR2. Активный центр состоит из активных дитиольных групп из RNR1, а также из диферрического центра и тирозильного радикала из субъединицы RNR2.
Другие остатки RNR2, такие как аспартат (D273), триптофан (W48) и тирозин (Y356), дополнительно стабилизируют тирозильный радикал в активном центре, тем самым обеспечивая перенос электронов. Эти остатки помогают переносить радикальный электрон от тирозина (Y122) из RNR2 к цистеину (C439) из RNR1. Перенос электрона начинается с тирозина RNR2 (Y122) и продолжается в RNR2 на триптофан (W48), который отделен от тирозина RNR1 (Y731) на 2,5 нанометра. Перенос электронов от RNR2 к RNR1 происходит через тирозин (от Y356 до Y731) и продолжается через тирозин (Y730) к цистеину (C439) в активном сайте. Сайт-ориентированные мутации первичной структуры RNR указывают на то, что все вышеперечисленные остатки участвуют в переносе свободного радикала на большие расстояния к активному сайту.
У комаров A. aegypti RNR1 сохраняет большинство важных аминокислотных остатков, включая аспартат (D64) и валин (V292 или V284), которые необходимы для аллостерической регуляции ; остатки пролина (P210 и P610), лейцина (L453 и L473) и метионина (M603), которые расположены в гидрофобном активном сайте; остатки цистеина (C225, C436 и C451), которые участвуют в удалении атома водорода и переносе радикального электрона в активном центре; остатки цистеина (C225 и C436), аспарагина (N434) и глутамата (E441), которые связывают рибонуклеотидный субстрат; остатки тирозина (Y723 и Y743), которые определяют перенос радикалов; и остатки цистеина (C838 и C841), которые используются для регенерации дитиоловых групп в активном центре.
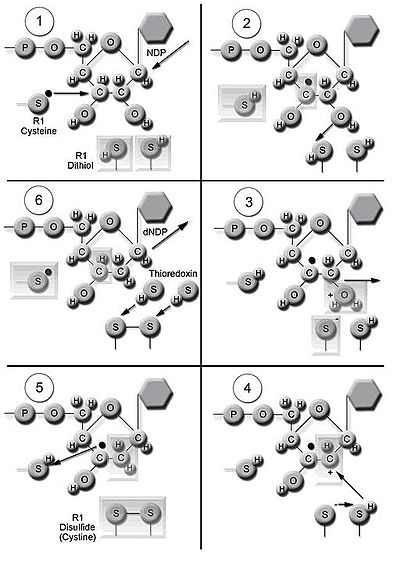 Механизм катализатора превращения рибонуклеотидов в дезоксирибонуклеотиды. (адаптировано из Nelson amp; Cox, 2000). (1) перенос электрона на субъединице RNR2 активирует остаток цистеина RNR1 в активном центре со свободным радикалом; (2) свободный радикал образует стабильный радикал на C-3, а радикал цистеина удаляет атом водорода; (3) катион образуется на C-2 путем переноса водорода из дитиольной группы и стабилизируется радикалом, что приводит к потере H2O из C-2; ( 4) водород переносится из дитиольной группы для восстановления катиона С-2; (5) радикал C-3 восстанавливается водородом, удаленным на стадии 2, и образуется тирозильный радикал; (6) редоксины переносят два водорода к дисульфидной группе, которая восстанавливает исходную конфигурацию.
Механизм катализатора превращения рибонуклеотидов в дезоксирибонуклеотиды. (адаптировано из Nelson amp; Cox, 2000). (1) перенос электрона на субъединице RNR2 активирует остаток цистеина RNR1 в активном центре со свободным радикалом; (2) свободный радикал образует стабильный радикал на C-3, а радикал цистеина удаляет атом водорода; (3) катион образуется на C-2 путем переноса водорода из дитиольной группы и стабилизируется радикалом, что приводит к потере H2O из C-2; ( 4) водород переносится из дитиольной группы для восстановления катиона С-2; (5) радикал C-3 восстанавливается водородом, удаленным на стадии 2, и образуется тирозильный радикал; (6) редоксины переносят два водорода к дисульфидной группе, которая восстанавливает исходную конфигурацию. Фермент рибонуклеотидредуктаза (RNR) катализирует синтез dNDP de novo. Катализ рибонуклеозид-5'-дифосфатов (NDP) включает восстановление у 2'-углерода рибозо-5-фосфата с образованием восстановленных 2'-дезоксипроизводным 2'-дезоксирибонуклеозид-5'-дифосфатов (dNDPs). Это снижение начинается с образования свободных радикалов. После однократного восстановления для RNR требуются электроны, пожертвованные дитиольными группами белка тиоредоксина. Регенерация тиоредоксина происходит, когда никотинамидадениндинуклеотидфосфат ( НАДФН ) обеспечивает два атома водорода, которые используются для восстановления дисульфидных групп тиоредоксина.
Три класса RNR имеют схожие механизмы восстановления NDP, но отличаются доменом, который генерирует свободный радикал, конкретным металлом в структуре металлопротеина и донорами электронов. Все классы используют свободнорадикальную химию. Редуктазы класса I используют центр железа с превращением двухвалентного железа в трехвалентное для образования свободного тирозильного радикала. Восстановление субстратов NDP происходит в аэробных условиях. Редуктазы класса I делятся на IA и IB из-за различий в регуляции. Редуктазы класса IA распространены у эукариот, эубактерий, бактериофагов и вирусов. Редуктазы класса IB обнаружены у эубактерий. Редуктазы класса IB могут также использовать радикал, образующийся при стабилизации биядерного марганцевого центра. Редуктазы класса II образуют свободный радикал 5'-дезоксиаденозил из кобаламина (кофермент B12) и имеют более простую структуру, чем редуктазы класса I и класса III. Восстановление NDP или рибонуклеотид-5'-трифосфатов (NTP) происходит либо в аэробных, либо в анаэробных условиях. Редуктазы класса II распространены в архебактериях, эубактериях и бактериофагах. Редуктазы класса III используют радикал глицина, образованный с помощью S-аденозилметионина и центра железа, серы. Снижение NTP ограничивается анаэробными условиями. Редуктазы класса III распространены в архебактериях, эубактериях и бактериофагах. Организмы не ограничиваются одним классом ферментов. Например, у E. coli есть RNR класса I и класса III.
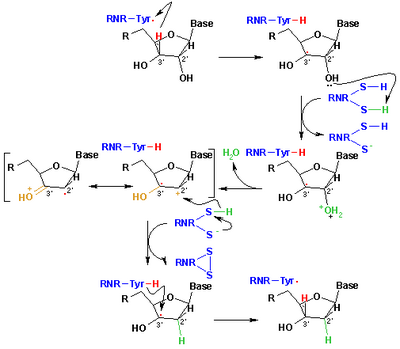 Механизм реакции РНР.
Механизм реакции РНР. Механизм, который в настоящее время принят для восстановления рибонуклеотидов до дезоксирибонуклеотидов, изображен на следующей схеме. Первый шаг включает отрыв 3'-H субстрата 1 радикалом Cys439. Впоследствии реакция включает отщепление одной молекулы воды от углерода C-2 'рибонуклеотида, катализируемое Cys225 и Glu441. На третьем этапе происходит перенос атома водорода от Cys225 на углерод C-2 '2'-кетильного радикала 3 после предыдущего переноса протона от Cys462 к Cys225. В конце этой стадии получают анион-радикальный дисульфидный мостик и промежуточный кетон 4 с закрытой оболочкой. Этот промежуточный продукт был идентифицирован во время превращения нескольких 2'-замещенных аналогов субстрата, а также с естественным субстратом, взаимодействующим с мутантами фермента. Следующим этапом является окисление анионного дисульфидного мостика с сопутствующим восстановлением субстрата с образованием 5. Спиновая плотность смещается от атомов серы к атому C-3 'субстрата с одновременным переносом протона от Glu441 на углерод C. -3 '. Последний этап является обратным первому этапу и включает перенос водорода от Cys439 к C-3 ', регенерируя исходный радикал и получая конечный продукт 6.
Теоретические модели некоторых этапов этих механизмов с использованием полной модели белка R1 можно найти в исследованиях, проведенных Cerqueira et al..
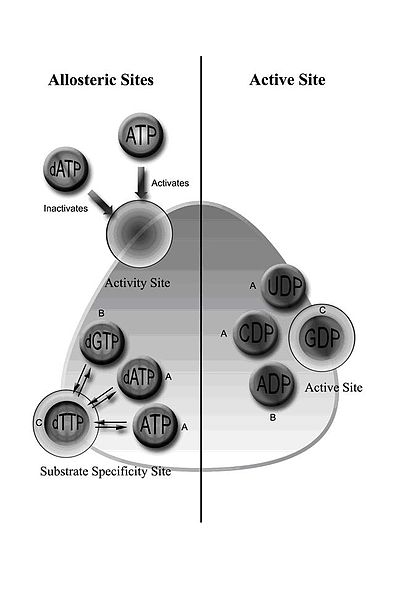 Регулирование класса I RNR. RNR класса I активируются путем связывания АТФ или инактивируются путем связывания dATP с сайтом активности, расположенным на субъединице RNR1. Когда фермент активируется, субстраты восстанавливаются, если соответствующие эффекторы связываются с сайтом специфичности аллостерического субстрата. A = когда dATP или ATP связаны в аллостерическом сайте, фермент принимает UDP и CDP в каталитический сайт; B = когда dGTP связан, ADP входит в каталитический сайт; C = когда dTTP связан, GDP входит в каталитический сайт. Субстраты (рибонуклеотиды UDP, CDP, ADP и GDP) превращаются в dNTP с помощью механизма, включающего образование свободных радикалов.
Регулирование класса I RNR. RNR класса I активируются путем связывания АТФ или инактивируются путем связывания dATP с сайтом активности, расположенным на субъединице RNR1. Когда фермент активируется, субстраты восстанавливаются, если соответствующие эффекторы связываются с сайтом специфичности аллостерического субстрата. A = когда dATP или ATP связаны в аллостерическом сайте, фермент принимает UDP и CDP в каталитический сайт; B = когда dGTP связан, ADP входит в каталитический сайт; C = когда dTTP связан, GDP входит в каталитический сайт. Субстраты (рибонуклеотиды UDP, CDP, ADP и GDP) превращаются в dNTP с помощью механизма, включающего образование свободных радикалов. RNR класса I включает субъединицы RNR1 и RNR2, которые могут связываться с образованием гетеродимерного тетрамера. RNR1 содержит оба аллостерических сайта, опосредуя регуляцию субстратной специфичности и активности. В зависимости от аллостерической конфигурации один из четырех рибонуклеотидов связывается с активным сайтом.
Регулирование RNR предназначено для поддержания сбалансированного количества dNTP. Связывание эффекторных молекул увеличивает или снижает активность РНР. Когда АТФ связывается с сайтом аллостерической активности, он активирует RNR. Напротив, когда dATP связывается с этим сайтом, он деактивирует RNR. Помимо контроля активности, аллостерический механизм также регулирует специфичность субстрата и гарантирует, что фермент продуцирует равное количество каждого dNTP для синтеза ДНК. Во всех классах связывание АТФ или дАТФ с аллостерическим сайтом индуцирует восстановление цитидин-5'-дифосфата (CDP) и уридин-5'-дифосфата (UDP); 2'-дезоксигуанозин-5'-трифосфат (dGTP) индуцирует восстановление аденозин-5'-дифосфата (ADP); и 2'-дезокситимидин-5'-трифосфат (dTTP) индуцирует восстановление гуанозин-5'-дифосфата (GDP) (Рисунок 1).
Редуктазы класса IB не ингибируются dATP, поскольку им не хватает примерно 50 N-концевых аминокислот, необходимых для сайта аллостерической активности. Кроме того, важно, чтобы активность рибонуклеотидредуктазы находилась под транскрипционным и посттранскрипционным контролем, поскольку синтез ДНК без повреждений зависит от сбалансированного пула дезоксирибонуклеотидов. Эукариотические клетки с редуктазами класса IA имеют механизм отрицательного контроля, чтобы отключить синтез dNTP по мере их накопления. Этот механизм защищает клетку от токсических и мутагенных эффектов, которые могут возникнуть в результате перепроизводства dNTP, поскольку изменения в сбалансированных пулах dNTP приводят к повреждению ДНК и гибели клеток. Хотя избыточная продукция dNTP или их несбалансированная поставка может привести к неправильному включению нуклеотидов в ДНК, поставка dNTPs может способствовать репарации ДНК. p53R2 представляет собой небольшую субъединицу рибонуклеотидредуктазы, которая может вызывать такую репарацию. Изменения в этом гомологе R2, индуцированном p53, могут вызывать истощение митохондриальной ДНК, и, следовательно, p53R2 служит основным фактором поставки dNTP.
RNR может использовать морфеиновую модель аллостерической регуляции.
Обычно ингибиторы RNR класса I можно разделить на три основные группы: ингибиторы трансляции, которые блокируют синтез фермента; ингибиторы димеризации, которые предотвращают ассоциацию двух субъединиц RNR (R1 и R2); и каталитические ингибиторы, которые инактивируют субъединицу R1 и / или субъединицу R2.
RNR класса I может ингибироваться пептидами, подобными C-концу RNR2. Эти пептиды могут конкурировать с RNR2 за связывание с RNR1, и в результате RNR1 не образует ферментативно активного комплекса с RNR2. Хотя C-конец белков RNR2 отличается у разных видов, RNR2 может взаимодействовать с RNR1 у разных видов. Когда C-конец RNR2 мыши был заменен на C-концевые (7 или 33) аминокислотные остатки RNR2 E. coli, химерная субъединица RNR2 все еще связывается с субъединицами RNR1 мыши. Однако им не хватает ферментативной активности, вероятно, из-за удаления остатков, участвующих в переносе свободнорадикального электрона от RNR2 к субъединице RNR1.
Небольшие пептиды могут специфически ингибировать связывание субъединиц RNR2 с RNR1, если они имеют значительное сходство с С-концом нормального RNR2. Это ингибирование связывания RNR2 с RNR1 было успешно протестировано на RNR вируса простого герпеса (HSV). Когда в конкурентных анализах использовали олигомер из 7 аминокислот (GAVVNDL), усеченный от С-конца субъединицы RNR2, это препятствовало образованию ферментативно активного комплекса с RNR1 нормальным RNR2. Другие малые пептидные ингибиторы, подобные С-концу RNR2, также успешно применялись для ингибирования ферментативной активности HSV RNR и, следовательно, репликации HSV. У мышей моделей стромального кератита и неоваскуляризация роговицы ( HSV глазной болезни ), небольшой RNR2 С-концевой аналоговый БИЛЬД +1263, как сообщались, ингибирует RNR и является эффективным в предотвращении этих заболеваний. В некоторых случаях, хотя лечение небольшими аналогами С-конца не может остановить распространение болезни, они все же могут помочь в выздоровлении. Сообщается, что в отношении устойчивого к ацикловиру HSV (PAAr5) небольшой пептидный ингибитор BILD 1633 в 5-10 раз более эффективен, чем BILD 1263, против кожной инфекции PAAr5. Комбинированная терапия (BILD 1633 и ацикловир) более эффективна для лечения местных повреждений у мышей. Эти данные предполагают, что небольшие пептидные ингибиторы, которые конкурируют с RNR2 за связывание с RNR1, полезны для предотвращения распространения HSV.
Галлий ингибирует RNR2, замещая Fe 3+ в активном центре. Мальтолат галлия представляет собой пероральную биодоступную форму галлия, которая использует эту ингибирующую активность для лечения рака, инфекций и других заболеваний.
Препараты гидроксимочевина и гадолиний Мотексафин препятствуют действию этого фермента.
 Кристаллографическая структура белка рибонуклеотидредуктазы R1E из
Кристаллографическая структура белка рибонуклеотидредуктазы R1E из 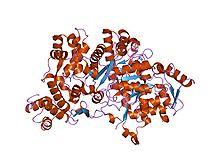 Структура белка рибонуклеотидредуктазы R1 (альфа-субъединица I класса).
Структура белка рибонуклеотидредуктазы R1 (альфа-субъединица I класса). Структура белка рибонуклеотидредуктазы R1E из Salmonella typhimurium.
Структура белка рибонуклеотидредуктазы R1E из Salmonella typhimurium. Структура белка рибонуклеотидредуктазы Escherichia coli R2.
Структура белка рибонуклеотидредуктазы Escherichia coli R2.