
Войти
| Птерозавры. Временной диапазон: Поздний триас - Поздний мел, 228–66 Ma До Ꞓ O S D C P T J K Pg N | |
|---|---|
 | |
| Копия скелетов Geosternbergia sternbergi, женские (слева) и мужские (справа) | |
| Научная классификация | |
| Царство: | Animalia |
| Тип: | Chordata |
| Clade: | Ornithodira |
| Clade: | †Pterosauromorpha. Padian, 1997 |
| Заказ: | †Pterosauria. Kaup, 1834 |
| Подгруппы | |
| |
 | |
| Распространение птерозавров ископаемых. Цветные названия видов или родов соответствуют их таксономической группе. | |
Птерозавры (; от греческого pteron и sauros, что означает «крылатая ящерица») летали рептилиями вымершей клады или отряда Pterosauria . Они существовали на протяжении большей части мезозоя : с конца триаса до конца мела (от 228 до 66 миллионов лет назад). Птерозавры - самые ранние позвоночные, которые, как известно, эволюционировали летающими двигателями. Их крылья были образованы мембраной из кожи, мышц и других тканей, простирающейся от лодыжек до резко удлиненного безымянного пальца.
Существовало два основных типа птерозавров. Базальные птерозавры (также называемые «нептеродактилоидными птерозаврами» или «рамфоринхоидами») были более мелкими животными с полностью зубчатыми челюстями и, как правило, длинными хвостами. Их широкие перепонки крыльев, вероятно, включали и соединяли задние лапы. На земле у них была бы неудобная поза, но их анатомия суставов и сильные когти сделали бы их эффективными альпинистами, и они, возможно, жили на деревьях. Базальные птерозавры были насекомоядными или хищниками мелких позвоночных. Позднее птерозавры (птеродактилоиды) приобрели разные размеры, формы и образы жизни. У птеродактилоидов были более узкие крылья со свободными задними конечностями, сильно редуцированные хвосты и длинные шеи с крупными головами. По земле птеродактилоиды хорошо ходили на всех четырех конечностях в вертикальном положении, стоя стопоход на задних лапах и сгибая крыловой палец вверх, чтобы ходить на трехпалой «руке». Они могли взлетать с земли, и следы окаменелостей показывают, что по крайней мере некоторые виды были способны бегать, переходить вброд или плавать. Их челюсти имели роговые клювы, а у некоторых групп не было зубов. У некоторых групп развились сложные головные гребни с половым диморфизмом.
Птерозавры носили слой волосоподобных волокон, известных как пикнофибры, которые покрывали их тела и части крыльев. Пикнофибры росли в нескольких формах, от простых нитей до ветвящихся пуховых перьев. Возможно, они гомологичны пуховым перьям, обнаруженным как у птичьих, так и у некоторых нептичьих динозавров, что позволяет предположить, что ранние перья эволюционировали у общего предка птерозавров и динозавров, возможно, как изоляция. При жизни птерозавры имели бы гладкую или пушистую шерсть, не похожую на птичьи перья. Это были теплокровные (эндотермические) активные животные. Дыхательная система имела эффективное однонаправленное «сквозное» дыхание с использованием воздушных мешков, которые в значительной степени выдавливали их кости. Птерозавры охватывали широкий диапазон взрослых размеров, от очень маленьких анурогнатид до самых крупных известных летающих существ всех времен, включая Quetzalcoatlus и Hatzegopteryx, достигшие размаха крыльев. не менее девяти метров. Сочетание эндотермии, хорошего снабжения кислородом и сильных мускулов сделало птерозавров сильными и способными летать.
популярные СМИ или широкая публика часто называют птерозавров "летающими динозаврами ", но динозавры определяются как потомки последнего общего предка Saurischia и Ornithischia, исключая птерозавров. Тем не менее птерозавры более тесно связаны с птицами и другими динозаврами, чем с крокодилами или другими живыми рептилиями, хотя они не являются предками птиц. Птерозавров также в просторечии называют птеродактилями, особенно в художественной литературе и журналистами. Однако технически птеродактиль относится только к членам рода Pterodactylus и, в более широком смысле, к членам подотряда Pterodactyloidea птерозавров.
Птерозавры имели множество разновидностей. образ жизни. Традиционно считается, что в эту группу едят рыбу, теперь в нее входят охотники на наземных животных, насекомоядные, плотоядные и даже хищники других птерозавров. Они размножаются яйцами, некоторые окаменелости которых были обнаружены.
Анатомия птерозавров сильно изменилась по сравнению с их рептильными предками приспособлением к полету. Кости птерозавров были полыми и заполненными воздухом, как у птиц. Это обеспечивало более высокую поверхность прикрепления мышцы для данного веса скелета. Костяные стенки часто были тонкими как бумага. У них была большая и килеватая грудина для мускулов полета и увеличенный мозг, способный координировать сложное летное поведение. Скелеты птерозавров часто обнаруживают значительное слияние. В черепе исчезли швы между элементами. У некоторых более поздних птерозавров позвоночник над плечами слился в структуру, известную как нотариум, которая служила для придания жесткости туловищу во время полета и обеспечения стабильной опоры для лопатки. Точно так же крестцовые позвонки могли образовывать единый синсакрум, в то время как кости таза также сливались.
Базальные птерозавры включают клады Dimorphodontidae (Dimorphodon ), Campylognathididae (Eudimorphodon, Campyognathoides ) и Rhamphorushynchidae (<281hynchidae)>, Скафогнат ).
Птеродактилоиды включают клады Ornithocheiroida (Istiodactylus, Ornithocheirus, Pteranodon ), Ctenochasmatoidea (Ctenochasma, Pterodactylus ), Dsungaripteroidea (Germanodactylus, Dsungaripterus ) и Azhdarchoidea (Tapejara, Tupuxuara, Quetzalco>).
Эти две группы перекрывались во времени, но самые ранние птерозавры в летописи окаменелостей - это базальные птерозавры, а новейшие птерозавры - птеродактилоиды.
Положение клады Anurognathidae (Anurognathus, Jeholopterus, Vesperopterylus ) обсуждается. Анурогнатиды (птерозавры с головой лягушки) были узкоспециализированными. Маленькие летуны с укороченными челюстями и широко разинутыми глазами, у некоторых были большие глаза, указывающие на ночной или крепскулярный образ жизни, щетину во рту и ступни, приспособленные к цеплянию. Параллельные адаптации наблюдаются у птиц и летучих мышей, охотящихся на насекомых в полете.
Птерозавры имели широкий диапазон размеров. В основном они были довольно крупными. Даже самые маленькие виды имели размах крыльев не менее 25 сантиметров (10 дюймов). Самые крупные формы представляют собой самых крупных из известных животных, когда-либо летающих, с размахом крыльев до 10–11 метров (33–36 футов).
Стоя, такие гиганты могут достигать роста современного жирафа.. Традиционно считалось, что птерозавры чрезвычайно легкие по сравнению со своими размерами. Позже стало понятно, что это будет означать нереально низкую плотность их мягких тканей. Поэтому некоторые современные оценки экстраполируют вес до 250 килограммов (550 фунтов) для самых крупных видов.
 Конический зуб, возможно, от Coloborhynchus
Конический зуб, возможно, от Coloborhynchus По сравнению с другие летающие группы позвоночных, птицы и летучие мыши, черепа птерозавров, как правило, были довольно большими. У большинства черепов птерозавров были удлиненные челюсти. Их кости черепа имеют тенденцию сращиваться у взрослых людей. Ранние птерозавры часто имели гетеродонтные зубы разного строения, а у некоторых все еще были зубы на нёбе. В более поздних группах зубы в основном приобрели коническую форму. Передние зубы часто были длиннее, образуя «захват для добычи» в поперечно расширенных кончиках челюстей, но размер и положение были очень разными у разных видов. С производным Pterodactyloidea черепа стали еще более вытянутыми, иногда превосходя по длине комбинированные шею и туловище. Это было вызвано растяжением и слиянием передней кости морды, предчелюстной кости, с костью верхней челюсти, верхней челюстью. В отличие от большинства архозавров, носовые и анторбитальные отверстия птеродактилоидных птерозавров слились в одно большое отверстие, называемое назоанторбитальным отверстием. Эта особенность, вероятно, эволюционировала, чтобы облегчить череп для полета. Напротив, кости за глазницей сжимались и вращались, сильно наклоняя заднюю часть черепа и выдвигая челюстной сустав вперед. черепная коробка была относительно большой для рептилий.
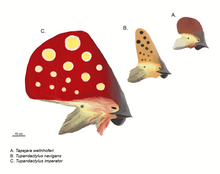 Реконструкция гребней: три гребенчатых ленточных червя. Сверху вниз: Tapejara wellnhoferi, Tupandactylus navigans, Tupandactylus imperator (в масштабе)
Реконструкция гребней: три гребенчатых ленточных червя. Сверху вниз: Tapejara wellnhoferi, Tupandactylus navigans, Tupandactylus imperator (в масштабе) В некоторых случаях окаменелые ороговевшие ткань клюва сохранилась, хотя у зубастых форм клюв небольшой, ограничивается кончиками челюстей и не затрагивает зубы. Некоторые развитые клювые формы были беззубыми, такие как Pteranodontidae и Azhdarchidae, и имели более крупные, протяженные и более похожие на птицу клювы. У некоторых групп были особые формы зубов. У Istiodactylidae были загнутые зубы для употребления в пищу мяса. Ctenochasmatidae использовали гребни с многочисленными игольчатыми зубьями для кормления с помощью фильтра; Птеродаустро мог иметь более тысячи щетиноподобных зубов. Dsungaripteridae покрывали свои зубы тканью челюстной кости для сокрушительной функции. Если зубы были, их помещали в отдельные лунки. Сменные зубы образовывались позади, а не под старыми зубами.
 Череп Thalassodromeus
Череп Thalassodromeus Общественный образ птерозавров определяется их сложными головными гребнями. На это повлиял характерный направленный назад гребень хорошо известного птеранодона. Основное положение таких гребней - это передняя часть морды, как вырост из предчелюстной кости, или задняя часть черепа как продолжение теменных костей, в этом случае это называется «надзатылочный гребень».. Передний и задний гребни могут присутствовать одновременно и могут быть слиты в единую большую структуру, наиболее обширная из которых представлена Tapejaridae. Никтозавр обладал причудливым рогом, похожим на гребень. Гребни были толщиной всего несколько миллиметров в поперечном направлении. Основание костного гребня обычно расширяется ороговевшей или другой мягкой тканью.
С 1990-х годов новые открытия и более тщательное изучение старых образцов показали, что гребни гораздо более распространены среди птерозавров, чем предполагалось ранее. То, что они были расширены или полностью состояли из кератина, который нелегко образуется в окаменелостях, вводило в заблуждение более ранние исследования. Для Pterorhynchus и Pterodactylus истинная протяженность этих гребней была обнаружена только с помощью ультрафиолетовой фотографии. В то время как ископаемые гребни обычно ограничивались более развитыми Pterodactyloidea, Pterorhynchus и Austriadactylus показывают, что даже некоторые ранние птерозавры обладали ими.
Как и верхняя челюсть, парные нижние челюсти птерозавров были очень хорошими. удлиненный. У продвинутых форм они, как правило, были короче верхнего черепа, потому что челюстной сустав находился в более переднем положении. Кости передней нижней челюсти, зубные кости или ossa dentalia, на конце были плотно сращены в центральный симфиз. Это заставило нижнюю челюсть функционировать как единое связное целое, нижней челюсти. Симфиз часто был очень тонким поперечно и длинным, составляя значительную часть длины челюсти, до 60%. Если бы на морде был гребень, симфиз мог бы иметь соответствующий гребень нижней челюсти, выступающий ниже. Зубчатые виды также имеют зубы в зубных челюстях. Нижняя челюсть открывается и закрывается простым вертикальным или «ортальным» движением вверх и вниз.
 шейный позвонок Arambourgiania
шейный позвонок Arambourgiania позвоночный столб птерозавров насчитывал от тридцати четырех до семидесяти позвонков. Позвонки перед хвостом были «проколозными»: гребешок (перед телом позвонка ) был вогнутым, и в него входило выпуклое продолжение в задней части предыдущего позвонка, мыщелок. Продвинутые птерозавры уникальны тем, что обладают особыми отростками, выступающими рядом с их мыщелком и мыщелком, экзапофизами, и на ягодицах также может быть небольшой зубец на средней линии, называемый гипапофизом.
 Шея Anhanguera была длиннее. чем туловище
Шея Anhanguera была длиннее. чем туловище Шея птерозавров была относительно длинной и прямой. У птеродактилоидов шея обычно длиннее туловища. Эта длина не вызвана увеличением количества позвонков, которое неизменно составляет семь. Некоторые исследователи включают два переходных «цервикодорсала», что доводит их число до девяти. Вместо этого сами позвонки стали более удлиненными, до восьми раз длиннее ширины. Тем не менее шейки матки были шире, чем высокие, что означало, что вертикальная подвижность шеи лучше, чем горизонтальная. Птеродактилоиды лишились всех шейных ребер. Шеи птерозавров, вероятно, были довольно толстыми и мускулистыми, особенно вертикально.
Туловище было относительно коротким и яйцевидным. Первоначально позвонков на спине птерозавров могло быть восемнадцать. У продвинутых видов все большее их количество имеет тенденцию встраиваться в крестец. Такие виды также часто демонстрируют слияние передних спинных позвонков в твердое целое, которое называется нотарием по схожей структуре у птиц. Это было приспособление, чтобы противостоять силам, вызванным взмахом крыльев. Нотариум включал от трех до семи позвонков, в зависимости от вида, а также от возраста человека. Эти позвонки могут быть соединены сухожилиями или слиянием их нервных отростков в «надраневральную пластинку». Их ребра тоже будут плотно впаяны в нотариальный зал. В целом ребра двуглавые. Крестец состоял из трех-десяти крестцовых позвонков. Их тоже можно было соединить через надбровную пластину, которая, однако, не касалась нотариума.
 Плечевой пояс, соединенный с нотарием
Плечевой пояс, соединенный с нотарием Хвосты птерозавров всегда были довольно тонкими. Это означает, что caudofemoralis втягивающая мышца, которая в большинстве базальных Archosauria обеспечивает главную движущую силу для задней конечности, была относительно неважной. Хвостовые позвонки амфикоелозные, тела позвонков на обоих концах вогнутые. Ранние виды имели длинные хвосты, содержащие до пятидесяти хвостовых позвонков, средние из которых были усилены удлиненными суставными отростками, зигапофизами и шевронами. Такие хвосты выполняли роль рулей, иногда заканчиваясь в задней части вертикальным ромбовидным или овальным флюгером. У птеродактилоидов хвосты были сильно уменьшены и никогда не были жесткими, у некоторых видов насчитывалось всего десять позвонков.
плечевой пояс представлял собой прочную структуру, которая передал силы махового полета на грудную клетку. Вероятно, он был покрыт толстыми слоями мышц. Верхняя кость, лопатка, была прямой перекладиной. Он был соединен с нижней костью, коракоидом, который у птерозавров относительно длинный. У продвинутых видов их совокупное целое, скапулокоракоид, ориентировано почти вертикально. В этом случае лопатка входила в выемку на стороне нотариуса, а коракоид также соединялся с грудиной. Таким образом, обе стороны вместе образуют жесткий замкнутый контур, способный выдерживать значительные силы. Особенностью было то, что грудные соединения коракоидов часто были асимметричными: один коракоид прикреплялся впереди другого. У продвинутых видов плечевой сустав переместился с лопатки на клювовидный. Шарнир имел седловидную форму и допускал значительный ход крыла. Он был обращен вбок и несколько вверх.
Грудь, образованная сросшейся парной грудиной, была широкой. У него был лишь неглубокий киль. Через грудные ребра он по бокам прикреплялся к спинным ребрам. Сзади имелся ряд брюшных ребер или gastralia, покрывающих весь живот. Спереди длинное острие, кристоспина, косо выступало вверх. Задний край грудины был самой глубокой точкой грудной клетки. Ключицы или межключицы полностью отсутствовали.
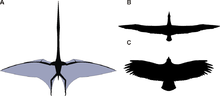 Реконструированная форма крыла в плане Quetzalcoatlus northropi (A) по сравнению с странствующим альбатросом Diomedea exulans (B) и Андский кондор Vultur gryphus (C). Они не в масштабе; размах крыльев Q. northropi, самого большого известного летающего животного, был более чем в три раза длиннее, чем у странствующего альбатроса.
Реконструированная форма крыла в плане Quetzalcoatlus northropi (A) по сравнению с странствующим альбатросом Diomedea exulans (B) и Андский кондор Vultur gryphus (C). Они не в масштабе; размах крыльев Q. northropi, самого большого известного летающего животного, был более чем в три раза длиннее, чем у странствующего альбатроса. Крылья птерозавра были образованы костями и оболочками кожи и других тканей. Первичные мембраны прикреплены к чрезвычайно длинному четвертому пальцу каждой руки и проходят по бокам тела. Вопрос о том, где они закончились, вызывает очень много споров, но с 1990-х годов была обнаружена дюжина образцов с сохранившимися мягкими тканями, которые, кажется, показывают, что они прикреплены к лодыжкам. Однако точная кривизна задней кромки все еще сомнительна.
 Некоторые экземпляры, такие как этот Rhamphorhynchus, сохраняют мембранную структуру
Некоторые экземпляры, такие как этот Rhamphorhynchus, сохраняют мембранную структуру Хотя исторически считались простыми кожистыми структурами, состоящими из кожи, Исследования с тех пор показали, что мембраны крыльев птерозавров представляют собой очень сложные динамические структуры, подходящие для активного стиля полета. Наружные крылья (от кончика до локтя) были усилены близко расположенными волокнами, называемыми актинофибриллами. Сами актинофибриллы состояли из трех отдельных слоев в крыле, которые при наложении друг на друга образовывали перекрестный узор. Функция актинофибрилл неизвестна, как и точный материал, из которого они были сделаны. В зависимости от их точного состава (кератин, мышцы, эластичные структуры и т. Д.) Они могли быть придающими жесткость или укрепляющими агентами во внешней части крыла. Мембраны крыльев также содержали тонкий слой мышечной, фиброзной ткани и уникальную сложную систему кровообращения, состоящую из петлевых кровеносных сосудов. Комбинация актинофибрилл и мышечных слоев, возможно, позволила животному регулировать расслабленность крыльев и выпуклость.
Как показывают полости в костях крыльев более крупных видов и мягкие ткани, сохранившиеся по крайней мере в одном экземпляре, некоторые птерозавры расширили свои система дыхательных воздушных мешков в мембрану крыла.
 Sordes, как показано здесь, свидетельствует о возможности того, что у птерозавров был круропатагиум - мембрана, соединяющая ноги, которые, в отличие от рукокрылых uropatagium, оставляют хвост свободным
Sordes, как показано здесь, свидетельствует о возможности того, что у птерозавров был круропатагиум - мембрана, соединяющая ноги, которые, в отличие от рукокрылых uropatagium, оставляют хвост свободным Мембрана крыла птерозавра разделена на три основных элемента. Первая, называемая пропатагиумом («передняя перепонка»), была самой передней частью крыла и прикреплялась между запястьем и плечом, создавая «переднюю кромку» во время полета. brachiopatagium («мембрана руки») был основным компонентом крыла, простирающимся от сильно удлиненного безымянного пальца руки до задних конечностей. Наконец, по крайней мере, у некоторых групп птерозавров была мембрана, которая протянулась между ног, возможно, соединялась с хвостом или включала его, называемая уропатагиум ; размер этой мембраны не определен, поскольку исследования Sordes, кажется, предполагают, что она просто соединяла ноги, но не затрагивала хвост (что делает его круропатагиумом ). Распространенное толкование состоит в том, что нептеродактилоидные птерозавры имели более широкий уро / круропатагий, протянутый между их длинными пятыми пальцами, а птеродактилоиды, лишенные таких пальцев, имели только мембраны, проходящие вдоль ног.>Среди палеонтологов были серьезные споры о том, прикрепляются ли основные перепонки крыльев (брахиопатагии) к задним конечностям, и если да, то где. Окаменелости рамфоринхоидов Sordes, анурогнатидов Jeholopterus и птеродактилоидов из формации Сантана, кажется, демонстрируют, что перепонка крыла действительно прикреплялась к задним конечностям, по крайней мере, у некоторых видов. Однако современные летучие мыши и белки-летяги демонстрируют значительные различия в протяженности мембран крыльев, и возможно, что, как и эти группы, разные виды птерозавров имели разный дизайн крыльев. Действительно, анализ пропорций конечностей птерозавра показывает, что были значительные различия, возможно, отражающие различные планы крыльев.
Костные элементы руки образовывали механизм, поддерживающий и расширяющий крыло. Рядом с телом плечевая кость или верхняя кость руки короткие, но мощно построенные. У него большой дельто-грудной гребень, к которому прикреплены основные летательные мышцы. Несмотря на приложенные к ней значительные силы, плечевая кость полая или пневматизированная внутри, усиленная костными распорками. Длинные кости нижней части руки, ulna и radius, намного длиннее плечевой кости. Вероятно, они были неспособны к пронации.
Кость, уникальная для птерозавров, известная как птероид, соединялась с запястьем и помогала поддерживать переднюю мембрану (пропатагиум) между запястьем и плечом. Свидетельства перепонки между тремя свободными пальцами передней конечности птерозавра предполагают, что эта передняя мембрана могла быть более обширной, чем простое соединение птероида с плечом, традиционно изображаемое при реставрации жизни. Положение самой птероидной кости было спорным. Некоторые ученые, в частности Мэтью Уилкинсон, утверждали, что птероид направлен вперед, расширяя переднюю мембрану и позволяя ей функционировать как регулируемый лоскут. Это мнение было опровергнуто в статье 2007 года Криса Беннета, который показал, что птероид не артикулируется, как считалось ранее, и не мог указывать вперед, а скорее был направлен внутрь к телу, как это традиционно интерпретируется. Дэвид Петерс в 2009 предположил, что птероид, сочлененный с «седлом» лучевой кости (проксимальный синкарпальный канал), и птероид и преаксиальный запястный сустав были перемещены в централи. Это подтверждается экземплярами Changchengopterus pani и Darwinopterus linglongtaensis, на которых птероид находится в сочленении с проксимальной синкарпальной костью.
Запястье птерозавра состоит из двух внутренних (проксимальных), сбоку от длинных костей руки) и четырех внешних (дистальных, сбоку кисти) запястья (кости запястья), за исключением птероидной кости, которая сама может быть модифицированной дистальной частью запястья. Проксимальные костей запястья сливаются вместе в «синкарпальные кости» у взрослых особей, в то время как три дистальных костей запястья сливаются, образуя дистальную синкарпальную кость. Оставшийся дистальный отдел запястья, называемый здесь медиальным запястным, но также называемый дистальным латеральным или преаксиальным запястным, сочленяется на удлиненной по вертикали двояковыпуклой фасетке на передней поверхности дистального отдела запястья. На медиальной части запястья имеется глубокая вогнутая ямка, которая открывается спереди, снизу и несколько медиально, внутри которой сочленяется птероид, согласно Уилкинсону.
У производных птеродактилоидов, таких как pteranodontians и azhdarchoids, пястные кости I-III маленькие и не соединяются с запястьями, а свисают в контакте с четвертой пястной костью. У этих производных видов четвертая пястная кость была чрезвычайно удлиненной, обычно равной или превышающей длину длинных костей нижней части руки. Пятая пястная кость была потеряна. У всех видов пальцы с первого по третий намного меньше четвертого, «крылатого пальца», и содержат две, три и четыре фаланги соответственно. Меньшие пальцы когтистые, размер ногтя варьируется у разных видов. У никтозаврид пальцы передних конечностей, кроме пальца крыла, полностью утрачены. На крылья приходится около половины или более общей длины крыла. Обычно он состоит из четырех фаланг. Их относительная длина, как правило, варьируется у разных видов, что часто используется для различения родственных форм. Четвертая фаланга обычно самая короткая. У него нет когтя, и он был полностью утерян никтозавридами. Он изогнут назад, что приводит к закругленной законцовке крыла, что снижает индуцированное лобовое сопротивление. Крыловой палец также несколько согнут вниз.
В положении стоя птерозавры, вероятно, опирались на пястные кости, при этом внешнее крыло загибалось назад. В этом положении «передние» стороны пястных костей повернуты назад. Это будет указывать меньшими пальцами наискось назад. По словам Беннета, это означало бы, что крыловой палец, способный описать самую большую дугу любого элемента крыла, до 175 °, был сложен не из-за сгибания, а из-за крайнего разгибания. Крыло автоматически складывалось при сгибании локтя.
 Ангангуэридный таз, вид сверху, с правой стороной, повернутой к наблюдателю
Ангангуэридный таз, вид сверху, с правой стороной, повернутой к наблюдателю таз птерозавров был среднего размера по сравнению с телом в целом. Часто сращивались три кости таза. подвздошная кость была длинной и низкой, ее передняя и задняя лопасти выступали горизонтально за края нижних костей таза. Несмотря на такую длину, стержневидная форма этих отростков указывает на то, что прикрепленные к ним мышцы задних конечностей были ограничены в силе. Узкая лобковая кость на виде сбоку слилась с широкой седалищной костью в седалищно-лобковую лопатку. Иногда сращивались и лопатки с обеих сторон, закрывая таз снизу и образуя тазовый канал. тазобедренный сустав не имеет перфорации и обеспечивает значительную подвижность ноги. Он был направлен под углом вверх, предотвращая идеально вертикальное положение ноги.
Передняя часть лобковых костей соединяется уникальной структурой - парными предлобными костями. Вместе они образовывали бугорок, покрывающий заднюю часть живота, между тазом и брюшными ребрами. Вертикальная подвижность этого элемента предполагает функцию дыхания, компенсирующую относительную жесткость грудной полости.
Задние конечности птерозавров были крепко сложены, но по сравнению с их размахом крыльев меньше, чем у птерозавров. птиц. Они были длинными по сравнению с длиной туловища. Бедренная кость была довольно прямой, голова составляла лишь небольшой угол с голенищем. Это означает, что ноги не держались вертикально ниже туловища, а были несколько раздвинуты. Большеберцовая кость часто срасталась с верхними костями лодыжки, образуя большеберцовую кость, которая была длиннее бедренной кости. Он может принимать вертикальное положение при ходьбе. Кость икроножной кости, как правило, была тонкой, особенно на ее нижнем конце, который у продвинутых форм не доходил до лодыжки, иногда уменьшая общую длину до трети. Обычно он сращивается с большеберцовой костью. Голеностопный сустав был простой, «мезотарзальной» петлей. Довольно длинная и тонкая плюсневая кость всегда была в той или иной степени растопыренной. Ступня была стопоходящей, что означало, что во время цикла ходьбы подошва плюсны была прижата к земле.
Между ранними птерозаврами и развитыми видами существовала явная разница в форме пятого пальца. Первоначально пятая плюсневая кость была крепкой и не очень укороченной. Он был соединен с лодыжкой в более высоком положении, чем другие плюсневые кости. У него был длинный и часто изогнутый подвижный пятый палец без когтей, состоящий из двух фаланг. Функция этого элемента была загадочной. Раньше считалось, что животные спят вверх ногами, как летучие мыши, свешиваясь с веток и используя пятые пальцы ног как крючки. Согласно другой гипотезе, они растягивали брахиопатагию, но в сочлененных окаменелостях пятые пальцы всегда согнуты к хвосту. Позже стало популярным предположение, что эти пальцы ног простирают между собой уропатагиум или круропатагиум. Поскольку пятые пальцы были на внешней стороне ступней, такая конфигурация была бы возможна только в том случае, если бы они поворачивали свои передние части в полете. Такое вращение могло быть вызвано отведением бедренной кости, что означало, что ноги будут раздвинуты. Это также вернет ступни в вертикальное положение. Затем они могли действовать как рули направления для контроля рыскания. У некоторых экземпляров между пальцами ног видны перепонки, позволяющие им выполнять функции управляющих поверхностей. (Cr) уроптагиум будет контролировать высоту звука. При ходьбе пальцы ног могут сгибаться вверх, чтобы оторвать мембрану от земли. У Pterodactyloidea пятая плюсневая кость была сильно редуцирована, а пятый палец, если он есть, был немного больше, чем окурок. Это говорит о том, что их мембраны были раздвоены, что увеличивало маневренность.
Пальцы с первого по четвертый были длинными. У них было две, три, четыре и пять фаланг соответственно. Часто третий палец ноги был самым длинным; иногда четвертый. Плоские суставы указывают на ограниченную подвижность. Эти пальцы ног были когтистыми, но когти были меньше, чем когти руки.
Редкие условия, которые позволили окаменеть останки птерозавров, иногда также сохраняли мягкие ткани. Современная синхротронная или ультрафиолетовая фотография выявила множество следов, невидимых невооруженным глазом. Их часто неточно называют «оттисками», но в основном они состоят из окаменелостей, естественных слепков и трансформаций исходного материала. Они могут включать роговые гребни, клювы или ножны когтей, а также различные лётные перепонки. В исключительном случае мышцы сохранились. Пятна на коже показывают небольшие круглые неперекрывающиеся чешуйки на подошвах стоп, лодыжках и концах пястных костей. Они покрыты подушечками, смягчающими удар при ходьбе. Чешуя на других частях тела не известна.
 Сордес сохранил пикнофибры
Сордес сохранил пикнофибры Большинство или все птерозавры имели волосы, -подобные волокна, известные как пикнофибры на голове и туловище. Термин «пикнофайбер», означающий «плотное волокно», был придуман палеонтологом Александром Келлнером и его коллегами в 2009 году. Пикнофибры были уникальными структурами, подобными, но не гомологичными (имеющими общее происхождение) с, волосами млекопитающих, примером конвергентной эволюции. Нечеткие покровы впервые были обнаружены у экземпляра Scaphognathus crassirostris в 1831 году, но вызвали серьезные сомнения. Начиная с 1990-х годов находки птерозавров и гистологические и ультрафиолетовые исследования образцов птерозавров неопровержимо доказывают: птерозавры были покрыты пикнофибрами. Sordes pilosus (что переводится как «волосатый демон») и Jeholopterus ninchengensis демонстрируют пикнофибры на голове и теле.
 Jeholopterus
Jeholopterus Присутствие пикнофибрилл строго указывает на то, что птерозавры были эндотермическими (теплокровными). Они способствовали терморегуляции, как это часто бывает у теплокровных животных, которым необходима изоляция для предотвращения чрезмерной потери тепла. Пикнофибры представляли собой гибкие короткие нити длиной около пяти-семи миллиметров, довольно простые по структуре с полым центральным каналом. Шкуры птерозавров могли быть сопоставимы по плотности со многими мезозойскими млекопитающими.
Нити птерозавров могли иметь общее происхождение с перьями, как предполагали в 2002 году Черкас и Джи. В 2009 году Келлнер пришел к выводу, что пикнофибры имеют структуру, аналогичную структуре теропод прото-перьев. Других это не убедило, считая разницу с «иглами», обнаруженными на многих образцах птицеподобных манирапторанов, слишком фундаментальной.
Исследование останков двух маленьких юрских времен в 2018 г. Птерозавры возраста из Внутренней Монголии, Китай обнаружили, что птерозавры имеют широкий спектр форм и структур пикнофибра, в отличие от однородных структур, которые, как обычно предполагалось, покрывают их. У некоторых из них были потертые концы, очень похожие по структуре на четыре разных типа перьев, известные у птиц или других динозавров, но почти никогда не известные у птерозавров до исследования, что предполагает гомологию. Ответ на это исследование был опубликован в 2020 году, где было высказано предположение, что структуры, наблюдаемые на анурогнатидах, на самом деле были результатом разложения актинофибрилл: типа волокна, используемого для усиления и придания жесткости крылу. Однако в ответ на это авторы статьи 2018 года указывают на то, что наличие структур распространяется за пределы патагия, а также на наличие актинофибрилл и нитей на Jeholopterus ningchengensis и Sordes pilosus. Различные формы филаментной структуры, присутствующие на анурогнатидах в исследовании 2018 года, также потребуют формы разложения, которая вызовет различные наблюдаемые `` филаментные '' формы. Таким образом, они пришли к выводу, что наиболее скупой интерпретацией структур является то, что они представляют собой нитчатые прото-перья.
 Гравировка оригинала Pterodactylus antiquus образец Эгида Верхелста II, 1784
Гравировка оригинала Pterodactylus antiquus образец Эгида Верхелста II, 1784 птерозавр окаменелости очень редки из-за их легкой костной конструкции. Полные скелеты, как правило, можно найти только в геологических слоях с исключительными условиями сохранности, так называемых Lagerstätten. Предметы одного такого лагерштетта, позднеюрский известняк Зольнхофена в Баварии, стали очень популярными среди богатых коллекционеров. В 1784 году итальянский натуралист Козимо Алессандро Коллини стал первым ученым в истории, описавшим окаменелость птерозавра. В то время концепции эволюции и вымирания были развиты недостаточно. Поэтому странное строение птерозавра шокировало, поскольку его нельзя было однозначно отнести к какой-либо существующей группе животных. Таким образом, открытие птерозавров сыграло бы важную роль в прогрессе современных приятелей. экологии и геологии. Если бы такие существа были все еще живы, то только море было бы надежной средой обитания, и Коллини предположил, что это могло быть плавающее животное, которое использовало свои длинные передние конечности как весла. Некоторые ученые продолжали поддерживать водную интерпретацию даже до 1830 года, когда немецкий зоолог Иоганн Георг Ваглер предположил, что Pterodactylus использовал свои крылья в качестве ласт и был связан с Ichthyosauria и Plesiosauria.
 Сумчатые птерозавры Ньюмана
Сумчатые птерозавры Ньюмана В 1800 году Иоганн Герман впервые предположил, что оно представляет собой летающее существо в письме к Жоржу Кювье. Кювье согласился в 1801 году, понимая, что это вымершая летающая рептилия. В 1809 году он придумал название Ptéro-Dactyle, «крылатый палец». В 1815 году он был латинизирован до Pterodactylus. Сначала большинство видов было отнесено к этому роду, и в конечном итоге термин «птеродактиль» был широко и неправильно применен ко всем членам Pterosauria. Сегодня палеонтологи ограничивают этот термин родом Pterodactylus или представителями Pterodactyloidea.
. В 1812 и 1817 гг. Самуэль Томас фон Зёммерринг переписал первоначальный образец и еще один. Он считал их связанными с птицами и летучими мышами. Хотя он ошибался в этом, его «модель летучей мыши» будет очень влиятельной в 19 веке. В 1843 году Эдвард Ньюман думал, что птерозавры летают сумчатыми. Поскольку модель летучей мыши правильно изображает птерозавров как покрытых шерстью и теплокровных, она лучше приближается к истинной физиологии птерозавров, чем «модель рептилий» Кювье. В 1834 году Иоганн Якоб Кауп ввел термин птерозаврия.
 Динамическая реконструкция диморфодона Сили
Динамическая реконструкция диморфодона Сили В 1828 году Мэри Эннинг в Англии нашла первый род птерозавров за пределами Германии, Ричард Оуэн названный Диморфодон, также первый известный нептеродактилоидный птерозавр. Позже в этом же веке в раннемеловом Cambridge Greensand были обнаружены тысячи окаменелостей птерозавров, которые, однако, были плохого качества и состояли в основном из сильно разрушенных фрагментов. Тем не менее, на основе этого будут названы многочисленные роды и виды. Многие из них были описаны Гарри Говьером Сили, в то время главным английским экспертом по этому вопросу, который также написал первую книгу о птерозаврах, Орнитозаврию, а в 1901 году - первую популярную книгу «Драконы воздуха». Сили считал птерозавров теплокровными и динамичными существами, близкими к птицам. Ранее эволюционист св. Джордж Джексон Миварт предположил, что птерозавры были прямыми предками птиц. Оуэн выступил против взглядов обоих мужчин, считая птерозавров хладнокровными «настоящими» рептилиями.
В США Отниэль Чарльз Марш в 1870 году обнаружил птеранодона в меле Ниобрара, тогда самый крупный из известных птерозавров, первый беззубый и первый из Америки. Эти слои также отображали тысячи окаменелостей, в том числе относительно полные скелеты, которые были сохранены в трехмерном виде вместо того, чтобы быть сильно сжатыми, как в случае с образцами из Зольнхофена. Это привело к лучшему пониманию многих анатомических деталей, таких как полость костей.
Между тем находки из Зольнхофена продолжались, составляя большинство обнаруженных полных образцов высокого качества. Они позволили идентифицировать большинство новых базальных таксонов, таких как Rhamphorhynchus, Scaphognathus и Dorygnathus. Этот материал положил начало немецкой школе исследований птерозавров, в которой летающие рептилии рассматривались как теплокровные, пушистые и активные мезозойские аналоги современных летучих мышей и птиц. В 1882 г. Марш и Карл Альфред Циттель опубликовали исследования оболочек крыльев у особей рамфоринхов. Немецкие исследования продолжались вплоть до 1930-х годов, описывая новые виды, такие как Anurognathus. В 1927 году Фердинанд Бройли обнаружил волосяные фолликулы в коже птерозавров, а палеоневролог Тилли Эдингер определили, что мозг птерозавров больше похож на мозг птиц, чем у современных хладнокровных. рептилии.
Напротив, английские и американские палеонтологи к середине двадцатого века в значительной степени потеряли интерес к птерозаврам. Они считали их неудачными эволюционными экспериментами, хладнокровными и чешуйчатыми, которые с трудом могли летать, а более крупные виды могли только парить, были вынуждены лазить по деревьям или бросаться со скал, чтобы взлететь. В 1914 году впервые была проведена количественная оценка аэродинамики птерозавров Эрнестом Хэнбери Хэнкином и, но они интерпретировали птеранодона как чистого планера. В течение 1940-х и 1950-х годов группа не исследовалась.
 Этот рисунок Чжэцзянгоптерус, сделанный Джоном Конвеем, иллюстрирует «новый облик» птерозавров
Этот рисунок Чжэцзянгоптерус, сделанный Джоном Конвеем, иллюстрирует «новый облик» птерозавров Ситуация для динозавров было сопоставимо. Начиная с 1960-х гг. Наступило ренессанс динозавров, быстрое увеличение количества исследований и критических идей, вызванное открытием дополнительных окаменелостей Дейнониха, чьи впечатляющие черты опровергли то, что стала укоренившейся ортодоксией. Аналогичным образом, в 1970 году описание пушистого птерозавра Сорд положило начало тому, что Роберт Баккер назвал возрождением птерозавров. В особенности Кевин Падиан пропагандировал новые взгляды, опубликовав серию исследований, изображающих птерозавров как теплокровных, активных и бегающих животных. Это совпало с возрождением немецкой школы благодаря работам Петера Велльнхофера, который в 1970-х заложил основы современной науки о птерозаврах. В 1978 году он опубликовал первый учебник по птерозаврам, Handbuch der Paläoherptologie, Teil 19: Pterosauria, а в 1991 году - вторую в истории научно-популярную книгу о птерозаврах, Энциклопедию птерозавров.
 Трехмерный сохраненный череп Анхангуэры santanae, из формации Santana, Brazil
Трехмерный сохраненный череп Анхангуэры santanae, из формации Santana, Brazil Это развитие ускорилось за счет эксплуатации двух новых Lagerstätten. В течение 1970-х годов в формации Сантана в раннем мелу в Бразилии начали образовываться меловые конкреции, которые, хотя часто ограничены по размеру и полноте содержащихся в них окаменелостей, прекрасно сохраняли трехмерные части скелета птерозавров. Немецкие и голландские институты купили такие конкреции у ископаемых браконьеров и приготовили их в Европе, что позволило их ученым описать множество новых видов и раскрыть целую новую фауну. Вскоре бразильские исследователи, в том числе Александр Келлнер, перехватили торговлю и назвали еще больше видов.
Еще более продуктивным был раннемеловой китайский Jehol Biota из Liaoning, который с 1990-х годов породил сотни прекрасно сохранившихся двумерных окаменелостей, часто демонстрирующих мягкие ткани. остается. Китайские исследователи, такие как Лю Цзюньчан, снова назвали много новых таксонов. Поскольку количество открытий увеличилось и в других частях мира, произошло внезапное увеличение числа названных родов. К 2009 году, когда их число увеличилось примерно до девяноста, этот рост не показал никаких признаков стабилизации. В 2013 году М. Виттон указал, что число обнаруженных видов птерозавров возросло до 130. Более девяноста процентов известных таксонов были названы во время «Возрождения». Многие из них были из групп, существование которых было неизвестно. Развитие вычислительной мощности позволило определить их сложные взаимосвязи с помощью количественного метода из кладистики. Новые и старые окаменелости дали гораздо больше информации, если подвергнуть их современной ультрафиолетовой или рентгеновской фотографии, или CAT-сканированию. К полученным данным были применены идеи из других областей биологии. Все это привело к значительному прогрессу в исследованиях птерозавров, в результате чего старые сообщения в научно-популярных книгах полностью устарели.
 Восстановление жизни Sharovipteryx, скользящего «проторозавра », который в некоторых противоречивых исследованиях считался близким родственник птерозавров.
Восстановление жизни Sharovipteryx, скользящего «проторозавра », который в некоторых противоречивых исследованиях считался близким родственник птерозавров. Поскольку анатомия птерозавра была настолько сильно изменена для полета, а непосредственные переходные ископаемые предшественники до сих пор не описаны, происхождение птерозавров полностью не изучено. Самые старые известные птерозавры уже были полностью адаптированы к летному образу жизни. Со времен Сили было признано, что птерозавры, вероятно, произошли от «архозавров», которых сегодня назвали бы архозавроморфами. В 1980-х годах ранний кладистический анализ показал, что они были авеметатарсалианцами (архозавры ближе к динозаврам, чем к крокодилам ). Поскольку это сделало бы их также довольно близкими родственниками динозавров, эти результаты были расценены Кевином Падианом как подтверждение его интерпретации птерозавров как двуногих теплокровных животных. Поскольку эти ранние анализы основывались на ограниченном количестве таксонов и признаков, их результаты были по своей сути неопределенными. Несколько влиятельных исследователей, которые отвергли выводы Падиана, предложили альтернативные гипотезы. предложил предков среди базальных архозавроморф, в частности длинношеих форм («проторозавров »), таких как танистрофеиды. Также было предложено размещение среди базальных архозавриформ, таких как Euparkeria. Некоторые базальные архозавроморфы на первый взгляд кажутся хорошими кандидатами в близких родственников птерозавров из-за их длинноногой анатомии; Одним из примеров является Sharovipteryx, «проторозавр» с кожными перепонками на задних конечностях, которые, вероятно, использовались для скольжения. Исследование 1999 г., проведенное Майклом Бентоном, показало, что птерозавры были авеметатарсалианами, тесно связанными с Scleromochlus, и назвало группу Ornithodira, чтобы включить птерозавров и динозавров.
 Восстановление жизни Scleromochlus, вероятно, ранний авеметатарсалий, обычно считающийся близким к птерозаврам.
Восстановление жизни Scleromochlus, вероятно, ранний авеметатарсалий, обычно считающийся близким к птерозаврам. Два исследователя, С. Кристофер Беннетт в 1996 году и палеохудожник Дэвид Питерс в 2000 году, опубликовали результаты анализа, в котором птерозавры были проторозаврами или были тесно связаны с их. Однако Петерс собрал новые анатомические данные, используя непроверенный метод под названием «Цифровая графическая сегрегация» (DGS), который включает в себя цифровое отслеживание изображений окаменелостей птерозавров с использованием программного обеспечения для редактирования фотографий. Беннетт восстановил птерозавров как близких родственников проторозавров после удаления характеристик задних конечностей из своего анализа, чтобы проверить возможность конвергентной эволюции на основе локомоций между птерозаврами и динозаврами. Ответ 2007 года Дэйва Хоуна и Майкла Бентона не смог воспроизвести этот результат, обнаружив, что птерозавры были тесно связаны с динозаврами даже без признаков задних конечностей. Они также критиковали Дэвида Питерса за то, что он делал выводы без доступа к первичным свидетельствам, то есть к самим окаменелостям птерозавров. Хоун и Бентон пришли к выводу, что, хотя для выяснения их взаимоотношений необходимо больше базальных птерозавроморфов, имеющиеся данные указывают на то, что птерозавры являются авеметатарсалианами, либо как родственная группа Scleromochlus, либо как ветвь между последним и Lagosuchus. Филогенетический анализ 2011 года, посвященный архозаврам, проведенный Стерлингом Несбиттом, основывался на гораздо большем количестве данных и обнаружил сильную поддержку птерозавров как авеметатарсалийцев, хотя Scleromochlus не был включен из-за его плохой сохранности. Исследование, посвященное архозавроморфам, проведенное Мартином Эскуррой в 2016 году, включало различных предполагаемых родственников птерозавров, но также обнаружило, что птерозавры ближе к динозаврам и не имеют отношения к большему количеству базальных таксонов. Основываясь на своем анализе 1996 года, Беннетт опубликовал исследование 2020 года по Scleromochlus, в котором утверждалось, что и Scleromochlus, и птерозавры не были архозавроморфами, не относящимися к архозаврам, хотя и не особенно тесно связаны друг с другом.
Связанная проблема. является источником полета птерозавров. Как и в случае с птицами, гипотезы можно разделить на два основных типа: «с нуля» или «с дерева». Подъем на дерево приведет к тому, что высота и сила тяжести будут обеспечивать как энергию, так и сильное давление выбора для начального полета. в 1983 году предложил гипотетический «проптерозавр»: древесное животное, похожее на ящерицу, развивающее перепонку между конечностями, сначала для безопасного парашюта, а затем, постепенно удлиняя безымянный палец, для скольжения. Однако последующие кладистические результаты не соответствовали этой модели. Ни протозавры, ни орнитодираны биологически не эквивалентны ящерицам. Кроме того, не совсем понятен переход между планирующим и махающим полетом. Более поздние исследования морфологии базальных задних конечностей птерозавров, кажется, подтверждают связь со Scleromochlus. Как и у этого архозавра, у базальных линий птерозавров есть стопоходящие задние конечности, которые демонстрируют адаптацию к сальтации.
 Скелет птерозавра, выставленный в Аризонском музее естественной истории в Столовой Аризоне.
Скелет птерозавра, выставленный в Аризонском музее естественной истории в Столовой Аризоне. Это было когда-то. считали, что конкуренция с ранними видами птиц могла привести к исчезновению многих птерозавров. Частично это связано с тем, что раньше считалось, что к концу мелового периода присутствовали только крупные виды птерозавров (теперь это неверно; см. Ниже). Считалось, что более мелкие виды вымерли, их ниша была заполнена птицами. Однако сокращение птерозавров (если оно действительно присутствует) не связано с разнообразием птиц, поскольку экологическое совпадение между двумя группами кажется минимальным. Фактически, по крайней мере, некоторые птичьи ниши были освоены птерозаврами до события KT. В конце мелового периода событие мелового – палеогенового вымирания, которое уничтожило всех нептичьих динозавров и большинство птичьих динозавров, а также многих других животных, похоже, также поглотило птерозавров.
В начале 2010-х годов было обнаружено несколько новых таксонов птерозавров, относящихся к кампану / маастрихту, например, орнитохейриды Пикси и «орнитохейрус », возможно птеранодонтиды и никтозавриды, несколько тейпежарид и неопределенный неаждархид Navajodactylus. Мелкие аждархоидные птерозавры также встречались в кампане. Это говорит о том, что позднемеловая фауна птерозавров была намного более разнообразной, чем считалось ранее, возможно, даже не уменьшившись значительно по сравнению с ранним меловым периодом.
Виды птерозавров мелких размеров, очевидно, присутствовали в формации Csehbánya, что указывает на более высокое разнообразие птерозавров позднего мела, чем считалось ранее. Недавние находки взрослого аждархида размером кошку также указывают на то, что маленькие птерозавры из позднего мелового периода могли просто редко сохраняться в летописи окаменелостей, чему способствовал тот факт, что существует сильное предубеждение против земных животных. позвоночные животные небольшого размера, такие как молодые динозавры, и что их разнообразие на самом деле могло быть намного больше, чем считалось ранее.
По крайней мере, некоторые нептеродактилоидные птерозавры дожили до позднего мелового периода, постулируя ситуацию таксонов Lazarus для позднемеловой фауны птерозавров.
В филогенетической таксономии клады Птерозаврию обычно определяют как узловую и прикрепляют к нескольким широко изученным таксонам, а также к тем, которые считаются примитивными. Одно исследование 2003 года определило Pterosauria как «самый недавний общий предок Anurognathidae, Preondactylus и Quetzalcoatlus и всех их потомков». Тем не менее, эти типы определения неизбежно оставят любые родственные виды, которые немного более примитивны, за пределами Pterosauria. Чтобы исправить это, было предложено новое определение, которое будет связывать название не с каким-либо конкретным видом, а с анатомической особенностью, наличием увеличенного безымянного пальца, поддерживающего перепонку крыла. Более широкая клада, Pterosauromorpha, была определена как все орнитодираны, более тесно связанные с птерозаврами, чем с динозаврами.
. Внутренняя классификация птерозавров имеет исторически было сложно, потому что в летописи окаменелостей было много пробелов. Начиная с 21 века, новые открытия заполняют эти пробелы и дают лучшее представление об эволюции птерозавров. Традиционно они были организованы в два подотряда : Rhamphorhynchoidea, «примитивная» группа длиннохвостых птерозавров, и Pterodactyloidea, «продвинутые» птерозавры с короткие хвосты. Однако от этого традиционного деления в значительной степени отказались. Rhamphorhynchoidea - это парафилетическая (неестественная) группа, поскольку птеродактилоиды произошли непосредственно от них, а не от общего предка, поэтому с увеличением использования кладистики она перестала пользоваться популярностью среди большинства ученых.
Точные отношения между птерозаврами до сих пор не установлены. Многие исследования взаимоотношений птерозавров в прошлом включали ограниченные данные и были весьма противоречивыми. Однако более новые исследования с использованием более крупных наборов данных начинают прояснять ситуацию. кладограмма (генеалогическое древо) ниже соответствует филогенетическому анализу, представленному Лонгричем, Мартиллом и Андресом в 2018 году.
 Диаграммы, показывающие дыхательное движение (две верхние) и систему внутреннего воздушного мешка (две нижние)
Диаграммы, показывающие дыхательное движение (две верхние) и систему внутреннего воздушного мешка (две нижние)  Адаптация полета птерозавра.
Адаптация полета птерозавра. Механика полета птерозавра не полностью изучена и не смоделирована. на этот раз.
Кацуфуми Сато, японский ученый, провел расчеты с использованием современных птиц и пришел к выводу, что птерозавр не может оставаться в воздухе. В книге «Осанка, передвижение и палеоэкология птерозавров» высказывается теория, что они могли летать благодаря богатой кислородом плотной атмосфере позднего мелового периода. Однако и Сато, и авторы книги «Осанка, передвижение и палеоэкология птерозавров» основывали свои исследования на устаревших теориях о том, что птерозавры похожи на морских птиц, и ограничение размера не распространяется на наземных птерозавров, таких как аждархиды и tapejarids. Кроме того, Даррен Нейш пришел к выводу, что атмосферные различия между настоящим и мезозойским периодом не нужны для гигантских размеров птерозавров.
 Реконструкция скелета четвероногого запускающего Pteranodon longiceps
Реконструкция скелета четвероногого запускающего Pteranodon longiceps Еще одна проблема, которая требует решения. Было трудно понять, как они сняли. Ранее предполагалось, что птерозавры в основном были хладнокровными летающими животными, которые, как современные ящерицы, получали тепло от окружающей среды, а не сжигали калории. В этом случае было неясно, как более крупные, огромных размеров, с неэффективным хладнокровным обменом веществ, могли управлять птичьим взлетом, используя только задние конечности для создания тяги для полета. Более поздние исследования показывают, что они теплокровны и обладают мощными летными мышцами и используют летательные мышцы для ходьбы на четвероногих. Марк Уиттон из Портсмутского университета и Майк Хабиб из Университет Джона Хопкинса предположил, что птерозавры использовали механизм прыжков для полета. Огромная сила их крылатых передних конечностей позволяла им легко взлетать. Поднявшись в воздух, птерозавры могли развивать скорость до 120 км / ч (75 миль в час) и преодолевать тысячи километров.
В 1985 году Смитсоновский институт поручил авиационному инженеру Полу Маккриди построить самолет. Полумасштабная рабочая модель Quetzalcoatlus northropi. Реплика запускалась с наземной лебедки. Он летал несколько раз в 1986 году и был снят как часть фильма Смитсоновского института IMAX На крыле.
Исследование 2009 года показало, что у птерозавров были легкие и воздушные мешочки. система и точно управляемый скелетный дыхательный насос, который поддерживает модель проточной легочной вентиляции у птерозавров, аналогичную модели птиц. Наличие системы подкожного воздушного мешка по крайней мере у некоторых птеродактилоидов привело бы к дальнейшему снижению плотности живого животного. Как и современные крокодилы, у птерозавров, по-видимому, был печеночный поршень, поскольку их плечево-грудные пояса были слишком негибкими, чтобы двигать грудиной, как у птиц, и они обладали сильной гастралией. Таким образом, их дыхательная система имела характеристики, сопоставимые с обеими современными кладами архозавров.
Рентгенологическое исследование полостей птерозавров мозга показало, что у животных (Rhamphorhynchus muensteri и Anhanguera santanae) были массивные флоккулы. flocculus - это область мозга, которая объединяет сигналы от суставов, мышц, кожи и органов равновесия. Флоккулы птерозавров занимали 7,5% общей массы мозга животных, больше, чем у любого другого позвоночного. Птицы имеют необычно большие флоккулы по сравнению с другими животными, но они занимают от 1 до 2% от общей массы мозга.
Флоккулы посылают нейронные сигналы, которые производят небольшие автоматические движения в мышцах глаза. Они удерживают изображение на сетчатке глаза животного. У птерозавров могли быть такие большие хлопья из-за их большого размера крыльев, что означало бы, что нужно было обработать гораздо больше сенсорной информации. Низкая относительная масса хлопьев у птиц также является результатом того, что у птиц в целом гораздо больший мозг; Хотя это считалось признаком того, что птерозавры жили в более простой по структуре окружающей среде или имели менее сложное поведение по сравнению с птицами, недавние исследования крокодилов и других рептилий показывают, что зауропсиды обычно достигают высокого уровня интеллекта с маленьким мозгом. Исследования эндокаста Аллкаруен показывают, что эволюция мозга у птеродактилоидов была модульным процессом.
 Вероятный след Аждархид ископаемое Haenamichnus uhangriensis.
Вероятный след Аждархид ископаемое Haenamichnus uhangriensis. Бедренные впадины птерозавров ориентированы немного вверх, а головка бедренной кости (бедренная кость) только умеренно обращена внутрь, что позволяет предположить, что птерозавры имели прямая стойка. Можно было бы во время полета поднять бедро в горизонтальное положение, как это делают планирующие ящерицы.
Было много споров о том, передвигались ли птерозавры как четвероногие или двуногие. В 1980-х годах палеонтолог Кевин Падиан предположил, что более мелкие птерозавры с более длинными задними конечностями, такие как диморфодон, могли ходить или даже бегать на двух ногах, помимо полета, как дорожные бегуны.. Однако позднее было обнаружено большое количество следов птерозавров с характерными четырехпалыми задними лапами и трехпалыми передними ногами; это безошибочные следы птерозавров, идущих на четвереньках.
Ископаемые следы показывают, что птерозавры стояли, соприкасаясь всей ступней с землей (стопоходящий ), подобно многим млекопитающим. как люди и медведи. Следы аждархид и нескольких неопознанных видов показывают, что птерозавры ходили в прямой позе с четырьмя конечностями, удерживаемыми почти вертикально под телом, - это энергосберегающая стойка, используемая большинством современных птиц и млекопитающих, а не распростертыми конечностями. современных рептилий. Действительно, прямостоячие конечности могут быть вездесущими у птерозавров.
 ископаемые следы показывают, что птерозавры, такие как Hatzegopteryx, были четвероногими и некоторыми довольно эффективными наземными хищниками.
ископаемые следы показывают, что птерозавры, такие как Hatzegopteryx, были четвероногими и некоторыми довольно эффективными наземными хищниками. Хотя изображались традиционно. анатомия некоторых птерозавров (особенно птеродактилоидов), столь же неуклюжих и неудобных на земле, предполагает, что они были умелыми ходунками и бегунами. Ранние птерозавры долгое время считались особенно громоздкими локомоторами из-за наличия больших круропатагий, но они, похоже, тоже в целом эффективны на земле.
Кости передних конечностей аждархид и орнитохейриды были необычно длинными по сравнению с другими птерозаврами, а у аждархид кости руки и кисти (пястные кости) были особенно удлиненными. Кроме того, в целом пропорции передних конечностей аждархид были аналогичны быстроходным копытным млекопитающим. Их задние конечности, с другой стороны, не были созданы для скорости, но они были длинными по сравнению с большинством птерозавров и позволяли большой шаг. Хотя аждархидные птерозавры, вероятно, не могли бегать, они были относительно быстрыми и энергоэффективными.
Относительный размер рук и ног у птерозавров (по сравнению с современными животными, такими как птицы) может указывать на тип образа жизни птерозавры вели по земле. Аждархидные птерозавры имели относительно маленькие ступни по сравнению с их размером тела и длиной ног, при этом длина ступни составляла всего около 25% -30% длины голени. Это говорит о том, что аждархиды лучше приспособились к ходьбе по сухой, относительно твердой земле. птеранодон имел немного большие ступни (47% длины большеберцовой кости ), тогда как у птерозавров, питающихся фильтром, таких как ктенохазматоиды, были очень большие ступни (69% большеберцовой кости). длина у Pterodactylus, 84% в Pterodaustro ), приспособленных к хождению по мягкой илистой почве, подобно современным болотным птицам. Хотя базальные птерозавры явно базируются на передних конечностях, задние конечности хорошо приспособлены к прыжкам, что указывает на связь с архозаврами, такими как Scleromochlus.
Следы, сделанные ктенохазматоидами, указывают на то, что эти птерозавры плавали с помощью задних конечностей. В целом у них большие задние лапы и длинные туловища, что указывает на то, что они, вероятно, были более приспособлены к плаванию, чем другие птерозавры. У птеранодонтов, напротив, есть несколько разновидностей плечевых суставов, которые интерпретируются как наводящие на мысль о водной версии типичного четвероногого катера, и несколько подобных бореоптерид должны были искать пищу во время плавания, поскольку они кажутся неспособными к фрегатным птицам. -подобный воздушный ястреб. Эти приспособления также наблюдаются у наземных птерозавров, таких как аждархид, которым, по-видимому, все еще нужно было запускать из воды, если они окажутся в ней. никтозаврид Альсионе может демонстрировать приспособления для ныряния с крылом, как современные олуши и тропические птицы.
Традиционно почти все птерозавры рассматривались как рыбоядные или рыбоядные, питающиеся на поверхности, и эта точка зрения все еще доминирует в научно-популярной науке. Сегодня считается, что многие группы птерозавров были наземными хищниками, всеядными или насекомоядными.
Ранее было признано, что маленькие Anurognathidae были ночными воздушными насекомоядными. Обладая очень гибкими суставами на крыловом пальце, широким треугольной формой крыла, большими глазами и коротким хвостом, эти птерозавры, вероятно, были аналогами козодоев или современных насекомоядных летучих мышей, будучи способными к высокой маневренности на относительно низких скоростях.
Интерпретации привычек базальных групп сильно изменились. Диморфодон, считавшийся в прошлом аналогом тупика, по строению челюсти, походке и плохим летным способностям обозначается как наземный / полуавтоматический хищник мелких млекопитающих, squamates и крупные насекомые. Его крепкие зубы заставляли Campylognathoides считаться универсальным или наземным хищником мелких позвоночных, но очень крепкая плечевая кость и морфология крыльев с высоким аспектом позволяют предположить, что он мог захватывать добычу за крыло. Маленькие насекомоядные Carniadactylus и более крупные Eudimorphodon были очень воздушными животными и быстрыми, подвижными летунами с длинными крепкими крыльями. Eudimorphodon был обнаружен с рыбными остатками в желудке, но его зубные ряды предполагают условную диету. Тонкокрылые Austriadactylus и Caviramus, вероятно, были наземными / полуодесными универсалистами. Кавирам, вероятно, обладал сильной силой укуса, что указывает на приспособление к твердой пище, которую можно было пережевывать из-за износа зубов.
Некоторые Rhamphorhynchidae, такие как Rhamphorhynchus сам или Доригнат, были рыбоедами с длинными тонкими крыльями, игольчатыми зубами и длинными тонкими челюстями. Sericipterus, Scaphognathus и Harpactognathus имели более крепкие челюсти и зубы (которые были зифодонтами в форме кинжала у Sericipterus) и более короткие и широкие крылья. Это были либо наземные / воздушные хищники позвоночных, либо врановые -подобные универсалы. Wukongopteridae подобные Darwinopterus впервые считались воздушными хищниками. Не имея прочной челюстной структуры или мощных летающих мускулов, они теперь считаются древесными или полуземными насекомоядными. В частности, Darwinopterus robustidens, по-видимому, был специалистом по жукам.
Среди птеродактилоидов наблюдается большее разнообразие в рационе. Pteranodontia содержал множество рыбоядных таксонов, таких как Ornithocheirae, Boreopteridae, Pteranodontidae и Nyctosauridae. Разделение ниши привело к тому, что орнитохейры и более поздние никтозавриды стали воздушными кормушками, такими как сегодняшние фрегаты (за исключением адаптированного для погружения Alcione elainus ), в то время как бореоптериды были пресноводными ныряющими животными, похожими на бакланов, и птеранодонтами-пелагическими ныряльщиками, похожими на олухи и олуши. Истиодактилиды, вероятно, были в основном падальщиками. Archaeopterodactyloidea добывали пищу в прибрежных или пресноводных местообитаниях. Germanodactylus и Pterodactylus были рыбоядными, а Ctenochasmatidae питались суспензией, использовав свои многочисленные мелкие зубы для фильтрации мелких организмов на мелководье. Pterodaustro был адаптирован для фламинго -подобного фильтрования.
Напротив, Azhdarchoidea в основном были наземными птерозаврами. Tapejaridae были древесными всеядными животными, дополнявшими семена и плоды мелкими насекомыми и позвоночными. Dsungaripteridae были специализированными моллюскоядными животными, использовавшими свои мощные челюсти, чтобы сокрушать раковины моллюсков и ракообразных. Thalassodromidae, вероятно, были наземными плотоядными животными. Thalassodromeus был назван в честь метода рыбной ловли, известного как «обезжиренное кормление», которое позже было признано биомеханически невозможным. Возможно, он преследовал относительно крупную добычу, учитывая усиленные челюстные суставы и относительно высокую силу укуса. Azhdarchidae теперь считается наземными хищниками, похожими на наземных птиц-носорогов или некоторых аистов., поедая любую добычу, которую они могли проглотить целиком. Хацегоптерикс был крепко сложенным хищником с относительно крупной добычей, включая динозавров среднего размера. Аланка, возможно, был моллюскоядным специалистом. Удивительное исследование было опубликовано в журнале Cretaceous Research. Палеонтологи анализировали челюсть одного вида птерозавров Lonchodraco giganteus и обнаружили отверстия около кончика его клюва, которые имели сходный рисунок, как у современных птиц, что указывает на тактильную привычку кормления у птерозавров.
Известно, что птерозавров съели тероподы. В выпуске Nature от 1 июля 2004 года палеонтолог Эрик Баффето обсуждает раннемеловые окаменелости трех шейных позвонков птерозавра со сломанным зубом спинозавр, скорее всего, Раздражитель, внедренный в него. Известно, что позвонки не были съедены и не подвергались перевариванию, поскольку суставы все еще сочленены.
 Ископаемые птеродактилоиды молодые особи из известняков Солнхофена
Ископаемые птеродактилоиды молодые особи из известняков Солнхофена Хотя очень мало известно о размножении птерозавров, считается, что, как и все динозавры, все птерозавры размножаются путем откладывания яиц, хотя такие находки очень редки. Первое известное яйцо птерозавра было найдено в карьерах Ляонина, в том же месте, где добывали пернатых динозавров. Яйцо было раздавлено, без признаков растрескивания, поэтому очевидно, что у яиц была кожистая скорлупа, как у современных ящериц. Это подтверждается описанием дополнительного яйца птерозавра, принадлежащего к роду Darwinopterus, описанного в 2011 году, которое также имело кожистую оболочку и, как и современные рептилии, но в отличие от птиц, было довольно маленьким по сравнению с размером матери. В 2014 году пять неуплюснутых яиц вида Hamipterus tianshanensis были обнаружены в раннемеловых отложениях на северо-западе Китая. Исследование скорлупы с помощью сканирующей электронной микроскопии показало наличие тонкого известкового слоя яичной скорлупы с мембраной под ним. Исследование структуры и химии яичной скорлупы птерозавров, опубликованное в 2007 году, показало, что птерозавры, вероятно, закопали свои яйца, как современные крокодилы и черепахи. Захоронение яиц было бы полезным для ранней эволюции птерозавров, поскольку оно позволяло бы больше приспособиться к снижению веса, но этот метод воспроизводства также ограничил бы разнообразие сред, в которых могли бы жить птерозавры, и, возможно, поставил бы их в невыгодное положение, когда они начали сталкиваться с экологической конкуренцией со стороны птиц.
Образец Darwinopterus демонстрирует, что, по крайней мере, у некоторых птерозавров была пара функциональных яичников, в отличие от одного функционального яичника у птиц, не считая уменьшения функциональные яичники как необходимое условие для полета.
Перепонки крыльев, сохранившиеся у эмбрионов птерозавров, хорошо развиты, что позволяет предположить, что птерозавры были готовы к полету вскоре после рождения. Однако томография сканирование окаменелых яиц Hamipterus предполагает, что у молодых птерозавров были хорошо развитые бедренные кости для ходьбы, но слабая грудь для полета. Неизвестно, верно ли это для других птерозавров. Были найдены окаменелости птерозавров возрастом от нескольких дней до недели (так называемые «хлопушки»), представляющие несколько семейств птерозавров, включая птеродактилид, рамфоринхид, ктенохасматид и аждарх. идентификаторы. У всех сохранились кости, которые показывают относительно высокую степень затвердевания (окостенения) для их возраста и пропорции крыльев, подобные взрослым. Фактически, многие взмахи птерозавров считались взрослыми особями и в прошлом помещались в отдельные виды. Кроме того, лоскуты обычно встречаются в тех же отложениях, что и взрослые особи и молодые особи одного и того же вида, такие как лоскуты Pterodactylus и Rhamphorhynchus, обнаруженные в известняках Solnhofen в Германии, и лоскуты Pterodaustro из Аргентины. Все они обитают в глубокой водной среде вдали от берега.
Для большинства видов птерозавров неизвестно, практиковали ли они какую-либо форму родительской заботы, но их способность летать, как только они вышли из яйца а многочисленные хлопушки, обнаруженные вдали от гнезд и рядом со взрослыми особями, привели большинство исследователей, в том числе Кристофера Беннета и Дэвида Анвина, к выводу, что детеныши зависели от своих родителей в течение относительно короткого периода времени, в период быстрого роста, в то время как крылья стали достаточно длинными, чтобы летать, а затем покинули гнездо, чтобы позаботиться о себе, возможно, в течение нескольких дней после вылупления. В качестве альтернативы они могли использовать сохраненные продукты из желтка для питания в течение первых нескольких дней жизни, как у современных рептилий, вместо того, чтобы зависеть от родителей в питании. Было показано, что в окаменелых гнездах Hamipterus сохраняются многие самцы и самки птерозавров вместе с их яйцами, как и в современных колониях морских птиц . Из-за того, что грудная клетка птенцов была недостаточно развита для полета, было высказано предположение, что Гамиптер, возможно, практиковал какую-то форму родительской заботы. Однако с тех пор это исследование подверглось критике. Большинство данных в настоящее время склоняется к тому, что вылупившиеся птерозавры суперпредколотные, аналогичные таковым у мегаподовых птиц, которые летают после вылупления без родительской заботы.
Скорость роста птерозавров после вылупления варьировалась в разных группах. У более примитивных длиннохвостых птерозавров («рамфоринхоидов »), таких как рамфоринхи, средняя скорость роста в течение первого года жизни составляла от 130% до 173%, что немного быстрее, чем скорость роста аллигаторы. После половой зрелости рост у этих видов замедлился, и рамфоринху потребовалось бы более трех лет, чтобы достичь максимального размера. Напротив, более продвинутые, крупные птеродактилоидные птерозавры, такие как Pteranodon, выросли до взрослых размеров в течение первого года жизни. Кроме того, птеродактилоиды имели определенный рост, что означало, что животные достигли фиксированного максимального размера взрослой особи и перестали расти.
Сравнение склеральных колец птерозавров и современные птицы и рептилии использовались, чтобы вывести характер повседневной активности птерозавров. Роды птерозавров Pterodactylus, Scaphognathus и Tupuxuara были выведены как дневные, Ctenochasma, Pterodaustro и Rhamphorhynchus были выведены как ночные, а Tapejara - как катемеральные, будучи активными в течение дня на короткие промежутки времени. В результате, возможно, питающиеся рыбой Ctenochasma и Rhamphorhynchus могли иметь сходные модели активности с современными ночными морскими птицами, а фильтрующие Pterodaustro могли иметь сходные модели активности с современными гусеобразными птицами, которые питаются ночью. Различия между моделями активности птерозавров Solnhofen Ctenochasma, Rhamphorhynchus, Scaphognathus и Pterodactylus также могут указывать на разделение ниш между этими родами.
 Quetzalcoatlus модели в Саут-Бэнк, созданные Марком Уиттоном к 350-летию Королевского общества
Quetzalcoatlus модели в Саут-Бэнк, созданные Марком Уиттоном к 350-летию Королевского общества Птерозавры были основным продуктом массовой культуры столько же, сколько и их кузены динозавры, хотя обычно они не фигурируют в фильмы, литература или другое искусство. Хотя изображение динозавров в популярных средствах массовой информации радикально изменилось в связи с достижениями палеонтологии, в основном устаревшее изображение птерозавров сохраняется с середины 20 века.
Для обозначения часто используется расплывчатый общий термин «птеродактиль». эти существа. Часто изображаемые животные представляют собой либо птеранодона, либо (нептеродактилоида) рамфоринха, либо их вымышленный гибрид. Многие детские игрушки и мультфильмы изображают «птеродактилей» с гребешками, подобными птеранодонам, и длинными, рамфоринхами, хвостами и зубами - комбинации, которой никогда не существовало в природе. Однако, по крайней мере, у одного птерозавра были и птеранодоноподобный гребень, и зубы: Ludodactylus, имя которого означает «игрушечный палец» из-за его сходства со старыми, неточными детскими игрушками. Птерозавров иногда неправильно идентифицировали как (предков) птиц, хотя птицы являются теропод динозаврами и не имеют близкого родства с птерозаврами.
Птерозавры использовались в художественной литературе в романе сэра Артура Конан Дойля Затерянный мир 1912 года и его экранизации 1925 года. С тех пор они появлялись в нескольких фильмах и телепрограммах, в том числе в фильме 1933 года Кинг-Конг и в фильме 1966 года Один миллион лет до нашей эры, в последнем, аниматор Рэй Харрихаузен Пришлось добавить неточные крылья летучей мыши к своим моделям покадровой анимации, чтобы мембраны не развалились, хотя именно эта ошибка была распространена в искусстве еще до того, как был снят фильм. Родан, вымышленный гигантский монстр (или кайдзю ), впервые появившийся в фильме 1956 года Родан, изображается как огромный облученный вид птеранодонов. Родан снимался в нескольких японских фильмах о Годзилле, выпущенных в 1960-х, 1970-х, 1990-х и 2000-х годах, а также появился в фильме американского производства 2019 года Годзилла: Король монстров.
После 1960-е годы птерозавры практически отсутствовали в заметных американских фильмах до 2001 года Парк Юрского периода III. Палеонтолог Дэйв Хоун отметил, что птерозавры в этом фильме не были значительно обновлены, чтобы отразить современные исследования. Сохраняющимися ошибками были зубы, в то время как беззубый птеранодон должен был изображаться, поведение гнездования, которое, как было известно, было неточным к 2001 году, и кожистые крылья, а не тугие мембраны мышечных волокон, необходимые для полета птерозавра.
В большинстве средств массовой информации Судя по всему, птерозавры изображены как рыбоядные, что не отражает их полное диетическое разнообразие. Их также часто изображают как воздушных хищников, похожих на хищных птиц, хватающих человеческие жертвы когтями на ногах. Однако известно, что только маленький анурогнатид Vesperopterylus обладает цепкими ногами; все другие птерозавры имеют плоские стопоходящие ступни без противопоставленных пальцев, и лапы обычно пропорционально малы, по крайней мере, в случае Pteranodontia.
| Викискладе есть медиафайлы, связанные с птерозаврией. |
| Викиисточником есть несколько оригинальных текстов, относящихся к: Птерозавры |