
Войти

Первичная структура белка представляет собой линейную последовательность аминокислот в пептиде или белок. По соглашению, первичная структура белка указывается, начиная с амино -конца (N) до карбоксильного -конца (C) конец. Биосинтез белка чаще всего осуществляется рибосомами в клетках. Пептиды также могут быть синтезированы в лаборатории. Первичные структуры белков могут быть непосредственно секвенированы или выведено из последовательностей ДНК.
Аминокислоты полимеризуются через пептидные связи с образованием длинный остов с выступающими вдоль него боковыми цепями различных аминокислот. В биологических системах белки продуцируются во время трансляции рибосомами клетки. Некоторые организмы могут также производить короткие пептиды посредством синтеза нерибосомных пептидов, которые часто используют аминокислоты, отличные от стандартных 20, и могут быть циклизованы, модифицированы и сшиты.
Пептиды можно синтезировать химическим с помощью ряда лабораторных методов. Химические методы обычно синтезируют пептиды в порядке, обратном (начиная с C-конца) биологическому синтезу белка (начиная с N-конца).
Последовательность белка обычно обозначается строкой букв, в которой перечислены аминокислоты, начиная с амино -концевого конца и заканчивая карбоксилом -терминальный конец. Можно использовать трехбуквенный или однобуквенный код для обозначения 20 встречающихся в природе аминокислот, а также смесей или неоднозначных аминокислот (аналогично нотации нуклеиновых кислот ).
Пептиды могут быть непосредственно секвенированы, или выведено из последовательностей ДНК. В настоящее время существуют большие базы данных последовательностей, которые сопоставляют известные белковые последовательности.
| Аминокислота | 3 буквы | 1 буква |
|---|---|---|
| аланин | Ala | A |
| аргинин | Arg | R |
| аспарагин | Asn | N |
| аспарагиновая кислота | Asp | D |
| цистеин | Cys | C |
| глутаминовая кислота | Glu | E |
| глутамин | Gln | Q |
| глицин | Gly | G |
| гистидин | His | H |
| изолейцин | Ile | I |
| лейцин | Leu | L |
| Lysine | Lys | K |
| Methionine | Met | M |
| Phenylalanine | Phe | F |
| Proline | Pro | P |
| Serine | Ser | S |
| Threonine | Thr | T |
| Триптофан | Trp | W |
| Тирозин | Tyr | Y |
| Валин | Val | V |
| Символ | Описание | Остатки р Представлено |
|---|---|---|
| X | Любая аминокислота или неизвестная | Все |
| B | Аспартат или аспарагин | D, N |
| Z | Глутамат или глутамин | E, Q |
| J | Лейцин или изолейцин | I, L |
| Φ | Гидрофобный | V, I, L, F, W, M |
| Ω | Ароматический | F, W, Y, H |
| Ψ | Алифатический | V, I, L, M |
| π | Маленький | P, G, A, S |
| ζ | Гидрофильный | S, T, H, N, Q, E, D, K, R, Y |
| + | Положительно заряженный | K, R, H |
| - | Отрицательно заряженный | D, E |
Как правило, полипептиды представляют собой неразветвленные полимеры, поэтому их первичная структура часто может быть заданные последовательностью аминокислот вдоль их основной цепи. Однако белки могут стать сшитыми, чаще всего с помощью дисульфидных связей, и первичная структура также требует указания сшивающих атомов, например, указания цистеинов, участвующих в дисульфиде белка. облигации. Другие поперечные связи включают десмозин.
. Хиральные центры полипептидной цепи могут подвергаться рацемизации. Хотя это не меняет последовательность, это влияет на химические свойства последовательности. В частности, L -аминокислоты, обычно присутствующие в белках, могут спонтанно изомеризоваться по атому 
Наконец, белок может претерпевать множество посттрансляционных модификаций, которые кратко описаны здесь.
N-концевая аминогруппа полипептида может быть модифицирована ковалентно, например,
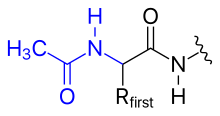 Фиг. 1 N-концевое ацетилирование
Фиг. 1 N-концевое ацетилирование 

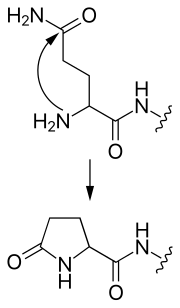 Рис. 2 Образование пироглутамата из N-концевого глутамина
Рис. 2 Образование пироглутамата из N-концевого глутамина 
С-концевая карбоксилатная группа полипептида также может быть модифицирована, например,
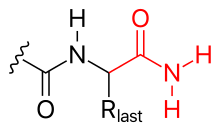 Рис. 3 С-концевое амидирование
Рис. 3 С-концевое амидирование Наконец, пептид боковые цепи также могут быть модифицированы ковалентно, например,
 атом. Несколько необычно, что эта модификация происходит в аппарате Гольджи, а не в эндоплазматическом ретикулуме. Подобно фосфорилированным тирозинам, сульфатированные тирозины используются для специфического распознавания, например, в рецепторах хемокинов на поверхности клетки. Как и при фосфорилировании, сульфатирование добавляет отрицательный заряд к ранее нейтральному сайту.
атом. Несколько необычно, что эта модификация происходит в аппарате Гольджи, а не в эндоплазматическом ретикулуме. Подобно фосфорилированным тирозинам, сульфатированные тирозины используются для специфического распознавания, например, в рецепторах хемокинов на поверхности клетки. Как и при фосфорилировании, сульфатирование добавляет отрицательный заряд к ранее нейтральному сайту.
 остатков цистеина для закрепления белков на клеточных мембранах. В отличие от GPI и миритоильных якорей, эти группы не обязательно добавляются на концах.
остатков цистеина для закрепления белков на клеточных мембранах. В отличие от GPI и миритоильных якорей, эти группы не обязательно добавляются на концах.Большинство модификаций полипептидов, перечисленных выше, происходят посттрансляционно, то есть после того, как белок был синтезирован на рибосоме, обычно встречающейся в эндоплазматическом ретикулуме, субклеточной органелле эукариотической клетки.
Многие другие химические реакции (например, цианилирование) были применены к белкам химиками, хотя они не обнаружены в биологических системах.
Помимо перечисленных выше, наиболее важной модификацией первичной структуры является расщепление пептида (химическим гидролизом или протеазы ). Белки часто синтезируются в неактивной форме-предшественнике; как правило, N-концевой или C-концевой сегмент блокирует активный сайт белка, ингибируя его функцию. Белок активируется отщеплением ингибирующего пептида.
Некоторые белки даже способны расщепляться. Как правило, гидроксильная группа серина (редко треонина) или тиольная группа остатка цистеина атакуют карбонильный углерод предыдущей пептидной связи, образуя тетраэдрически связанный промежуточный продукт [классифицируемый как гидроксиоксазолидин (Ser / Thr) или гидрокситиазолидин ( Cys) промежуточный]. Этот промежуточный продукт имеет тенденцию превращаться в амидную форму, вытесняя атакующую группу, поскольку амидной форме обычно способствует свободная энергия (предположительно из-за сильной резонансной стабилизации пептидной группы). Однако дополнительные молекулярные взаимодействия могут сделать амидную форму менее стабильной; вместо этого удаляется аминогруппа, что приводит к образованию сложноэфирной (Ser / Thr) или тиоэфирной (Cys) связи вместо пептидной связи. Эта химическая реакция называется.
Связь сложный эфир / тиоэфир может быть разделена несколькими способами:
Предположение, что белки представляют собой линейные цепи α-аминокислот, было сделано почти одновременно двумя учеными на той же конференции в 1902 году, 74-м заседании Общество немецких ученых и врачей, проходившее в Карловых Варах. Франц Хофмайстер сделал предложение утром, основываясь на своих наблюдениях за реакцией биурета в белках. Через несколько часов за Хофмайстером последовал Эмиль Фишер, который накопил множество химических деталей, подтверждающих модель пептидной связи. Для полноты предположение, что белки содержат амидные связи, было сделано еще в 1882 г. французским химиком Э. Гримо.
Несмотря на эти данные и более поздние доказательства того, что протеолитически расщепленные белки давали только олигопептиды, идея о том, что белки были линейными, неразветвленные полимеры аминокислот не сразу приняли. Некоторые уважаемые ученые, такие как Уильям Эстбери сомневались, что ковалентные связи были достаточно прочными, чтобы удерживать вместе такие длинные молекулы; они опасались, что тепловое возбуждение может расколоть такие длинные молекулы. Герман Штаудингер столкнулся с аналогичными предрассудками в 1920-х годах, когда он утверждал, что каучук состоит из макромолекул.
Таким образом, возникло несколько альтернативных гипотез. Гипотеза коллоидного белка утверждала, что белки представляют собой коллоидные сборки более мелких молекул. Эта гипотеза была опровергнута в 1920-х годах измерениями ультрацентрифугирования, проведенными Теодором Сведбергом, которые показали, что белки имеют четко определенную воспроизводимую молекулярную массу, и с помощью электрофоретических измерений, проведенных Арне Тизелиус, которые показали, что белки были одиночные молекулы. Вторая гипотеза, гипотеза циклола, выдвинутая Дороти Ринч, предполагала, что линейный полипептид претерпел химическую циклоловую перегруппировку C = O + HN 
Можно сказать, что любой гетерополимер с линейной цепью имеет «первичную структуру» по аналогии с использованием термина для белков, но это использование редко по сравнению с чрезвычайно распространенное использование по отношению к белкам. В РНК, которая также имеет обширную вторичную структуру, линейная цепь оснований обычно называется просто «последовательностью», как в ДНК (которая обычно образует линейную двойную спираль с небольшой вторичной структурой). Другие биологические полимеры, такие как полисахариды, также могут считаться имеющими первичную структуру, хотя их использование не является стандартным.
Первичная структура биологического полимера в значительной степени определяет трехмерную форму (третичная структура ). Последовательность белка может использоваться для прогнозирования локальных особенностей, таких как сегменты вторичной структуры или трансмембранные области. Однако сложность сворачивания белка в настоящее время не позволяет предсказывать третичную структуру белка только по его последовательности. Знание структуры подобной гомологичной последовательности (например, члена того же семейства белков ) позволяет с высокой точностью предсказывать третичную структуру по гомологии моделирование. Если доступна полноразмерная белковая последовательность, можно оценить ее общие биофизические свойства, такие как его изоэлектрическая точка.
Семейства последовательностей часто определяются с помощью кластеризации последовательностей и проекты структурной геномики направлены на создание набора репрезентативных структур для покрытия пространства последовательностей возможных неизбыточных последовательностей.