
Войти
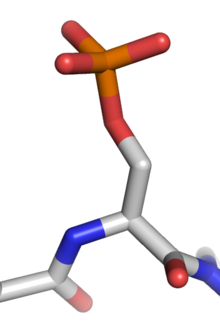 Модель фосфорилированного остатка серина
Модель фосфорилированного остатка серина  серина в аминокислотной цепи до и после фосфорилирования.
серина в аминокислотной цепи до и после фосфорилирования. Фосфорилирование белка представляет собой обратимую посттрансляционную модификацию белков, в которой аминокислотный остаток фосфорилируется протеин киназой путем добавления ковалентно связанной фосфатной группы. Фосфорилирование изменяет структурную конформацию белка, заставляя его активироваться, дезактивироваться или изменять его функцию. Приблизительно 13000 белков человека имеют участки, которые фосфорилируются.
Обратная реакция фосфорилирования называется дефосфорилированием и катализируется белковыми фосфатазами. Протеинкиназы и фосфатазы работают независимо и сбалансированно, регулируя функцию белков.
Наиболее часто фосфорилируемыми аминокислотами являются серин, треонин, тирозин у эукариот, а также гистидин у прокариот и растений (хотя теперь известно, что он часто встречается у людей). Эти фосфорилирования играют важную и хорошо изученную роль в сигнальных путях и метаболизме. Однако другие аминокислоты также могут фосфорилироваться посттрансляционно, в том числе аргинин, лизин, аспарагиновая кислота, глутаминовая кислота и цистеин, и недавно было обнаружено, что эти фосфорилированные аминокислоты присутствуют в экстрактах клеток человека и фиксированных клетках человека, с помощью комбинации анализа на основе антител (для pHis) и масс-спектрометрии (для всех других аминокислот).
О фосфорилировании белков впервые сообщил в 1906 году Фебус Левен из Института медицинских исследований Рокфеллера с открытием фосфорилированного вителлина. Однако прошло почти 50 лет до открытия ферментативного фосфорилирования белков протеинкиназами.
В 1906 г. Phoebus Levene в Институте медицинских исследований Рокфеллера идентифицировали фосфат в белке вителлин (фосвитин), а к 1933 году обнаружил фосфосерин. в казеине, совместно с Фрицем Липманном. Однако прошло еще 20 лет, прежде чем Юджин П. Кеннеди описал первое «ферментативное фосфорилирование белков». Первый фермент фосфорилаза был открыт Карлом и Герти Кори в конце 1930-х годов. Карл и Герти Кори обнаружили две формы гликогенфосфорилазы, которые они назвали A и B, но не правильно поняли механизм преобразования формы B в форму A. Взаимопревращение фосфорилазы b в фосфорилазу a было позже описано Эдмондом Фишером и Эдвином Кребсом, а также Wosilait и Sutherland, включая механизм фосфорилирования / дефосфорилирования.. Было обнаружено, что фермент, названный киназа фосфорилазы и Mg-АТФ, необходим для фосфорилирования гликогенфосфорилазы путем содействия переносу γ-фосфорильной группы АТФ к остатку серина на фосфорилазе b. Протеинфосфатаза 1 способна катализировать дефосфорилирование фосфорилированных ферментов за счет удаления фосфатной группы. Эрл Сазерленд объяснил в 1950 году, что активность фосфорилазы увеличивалась и, таким образом, стимулировался гликогенолиз, когда срезы печени инкубировали с адреналином и глюкагоном. Фосфорилирование считалось специфическим механизмом контроля одного метаболического пути до 1970-х годов, когда Лестер Рид обнаружил, что комплекс митохондриальной пируватдегидрогеназы инактивирован фосфорилированием. Также в 1970-х годах термин «мультисайтовое фосфорилирование» был придуман в ответ на открытие белков, которые фосфорилируются по двум или более остаткам двумя или более киназами. В 1975 году было показано, что цАМФ-зависимые протеинкиназы фосфорилируют сериновые остатки на определенных мотивах аминокислотной последовательности. Рэй Эриксон обнаружил, что v-Src является киназой, а Тони Хантер обнаружил, что v-Src фосфорилирует остатки тирозина на белках в 1970-х годах. В начале 1980 года была определена аминокислотная последовательность первой протеинкиназы, которая помогла генетикам понять функции регуляторных генов. В конце 1980-х и начале 1990-х годов была очищена первая протеинтирозинфосфатаза (PTP1B), и было совершено открытие, а также клонирование киназ JAK, что привело к многим научное сообщество назвало 1990-е десятилетием каскадов протеинкиназ. Эдмонд Фишер и Эдвин Кребс были удостоены Нобелевской премии в 1992 году «за открытия, касающиеся обратимого фосфорилирования белков как биологического механизма регуляции».
Фосфорилирование вводит заряженную и гидрофильную группу в боковую цепь аминокислот, возможно, изменяя структуру белка, изменяя взаимодействия с соседними аминокислотами. Некоторые белки, такие как p53, содержат несколько сайтов фосфорилирования, облегчая сложную многоуровневую регуляцию. Из-за легкости, с которой белки могут быть фосфорилированы и дефосфорилированы, этот тип модификации представляет собой гибкий механизм, позволяющий клеткам реагировать на внешние сигналы и условия окружающей среды.
Обратимое фосфорилирование белков происходит как в прокариотах и эукариотические организмы. Подсчитано, что у человека, мыши и дрожжей существует 230 000, 156 000 и 40 000 сайтов фосфорилирования соответственно. киназы фосфорилируют белки, а фосфатазы дефосфорилируют белки. Многие ферменты и рецепторы «включаются» или «выключаются» за счет фосфорилирования и дефосфорилирования. Обратимое фосфорилирование приводит к конформационному изменению в структуре многих ферментов и рецепторов, заставляя их активироваться или дезактивироваться. Фосфорилирование обычно происходит по остаткам серина, треонина, тирозина и гистидина в эукариотических белках. Фосфорилирование эукариотических белков гистидином происходит гораздо чаще, чем фосфорилирование тирозина. В прокариотических белках фосфорилирование происходит по остаткам серина, треонина, тирозина, гистидина или аргинина или лизина. Добавление молекулы фосфата (PO 4) к неполярной R-группе аминокислотного остатка может превратить гидрофобную часть белка в полярную и чрезвычайно гидрофильную часть молекулы. Таким образом, динамика белка может вызывать конформационные изменения в структуре белка через дальнодействующую аллостерию с другими гидрофобными и гидрофильными остатками в белке.
Одним из таких примеров регуляторной роли, которую играет фосфорилирование, является белок-супрессор опухоли p53. Белок p53 сильно регулируется и содержит более 18 различных сайтов фосфорилирования. Активация р53 может привести к остановке клеточного цикла, который может быть обращен вспять при некоторых обстоятельствах, или к апоптотической гибели клеток. Эта активность происходит только в ситуациях, когда клетка повреждена или физиология нарушена у нормальных здоровых людей.
По сигналу деактивации белок снова становится дефосфорилированным и перестает работать. Это механизм во многих формах передачи сигнала, например, способ, которым поступающий свет обрабатывается в светочувствительных клетках сетчатки.
Регуляторные роли фосфорилирования включают:
Выявление событий фосфорилирования комплекса сигнального пути может быть затруднено. В клеточных сигнальных путях белок A фосфорилирует белок B, а B фосфорилирует C. Однако в другом сигнальном пути белок D фосфорилирует A или фосфорилирует белок C. Глобальные подходы, такие как фосфопротеомика, исследование фосфорилированных белков, которое является подветвью протеомики, в сочетании с протеомикой на основе масс-спектрометрии, было использовано для выявления и количественной оценки динамических изменений фосфорилированных белков с течением времени.. Эти методы становятся все более важными для систематического анализа сложных сетей фосфорилирования. Они были успешно использованы для определения динамических изменений статуса фосфорилирования более чем 6000 сайтов после стимуляции эпидермальным фактором роста. Другой подход к пониманию сети фосфорилирования заключается в измерении генетических взаимодействий между множественными фосфорилирующими белками и их мишенями. Это выявляет интересные повторяющиеся паттерны взаимодействий - сетевые мотивы. Вычислительные методы были разработаны для моделирования сетей фосфорилирования и прогнозирования их ответов при различных возмущениях.
Эукариотическая ДНК организована с гистоновыми белками в определенные комплексы, называемые хроматином. Структура хроматина функционирует и облегчает упаковку, организацию и распространение эукариотической ДНК. Однако он оказывает негативное влияние на несколько фундаментальных биологических процессов, таких как транскрипция, репликация и репарация ДНК, ограничивая доступность определенных ферментов и белков. Было показано, что посттрансляционная модификация гистонов, такая как фосфорилирование гистонов, изменяет структуру хроматина путем изменения взаимодействий белок: ДНК или белок: белок. Посттрансляционные модификации гистонов изменяют структуру хроматина. Наиболее часто ассоциированное фосфорилирование гистонов происходит во время клеточных ответов на повреждение ДНК, когда фосфорилированный гистон H2A разделяет большие домены хроматина вокруг места разрыва ДНК. Исследователи выяснили, влияют ли модификации гистонов напрямую на транскрипцию, управляемую РНК-полимеразой II. Исследователи выбирают белки, которые, как известно, модифицируют гистоны, чтобы проверить их влияние на транскрипцию, и обнаружили, что индуцированная стрессом киназа, MSK1, ингибирует синтез РНК. Ингибирование транскрипции с помощью MSK1 было наиболее чувствительным, когда матрица находилась в хроматине, поскольку матрицы ДНК не в хроматине были устойчивы к эффектам MSK1. Было показано, что MSK1 фосфорилирует гистон H2A по серину 1, а мутация серина 1 в аланин блокирует ингибирование транскрипции с помощью MSK1. Таким образом, результаты показали, что ацетилирование гистонов может стимулировать транскрипцию путем подавления ингибирующего фосфорилирования киназой, такой как MSK1.
Внутри белка фосфорилирование может происходить по нескольким аминокислотам. Считается, что наиболее распространенным является фосфорилирование серина, за которым следует треонин. Фосфорилирование тирозина встречается относительно редко, но оно стоит во главе многих сигнальных путей фосфорилирования белков (например, в рецепторах, связанных с тирозинкиназой) у большинства эукариот. Фосфорилирование аминокислот, таких как серин, треонин и тирозин, приводит к образованию фосфопротеина, когда фосфатная группа фосфопротеина реагирует с группой -ОН боковой цепи Ser, Thr или Tyr в реакция этерификации. Однако, поскольку белки, фосфорилированные тирозином, относительно легко очистить с использованием антител, сайты фосфорилирования тирозина относительно хорошо изучены. Гистидин и аспартат фосфорилирование происходит у прокариот как часть двухкомпонентной передачи сигналов и в некоторых случаях у эукариот в некоторых путях передачи сигнала. Анализ фосфорилированного гистидина с использованием стандартных биохимических и масс-спектрометрических подходов намного сложнее, чем анализ Ser, Thr или Tyr. и У прокариот, архей и некоторых низших эукариот азот гистидина действует как нуклеофил и связывается с фосфатной группой. После фосфорилирования гистидина регуляторный домен регулятора ответа катализирует перенос фосфата в аспартат.
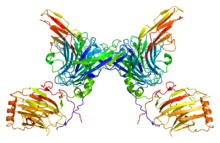 Рецепторная тирозинкиназа AXL, демонстрирующая симметрию димеризованных рецепторов
Рецепторная тирозинкиназа AXL, демонстрирующая симметрию димеризованных рецепторов Хотя фосфорилирование тирозина обнаруживается в относительно низком количестве, оно хорошо изучено из-за простоты очистки фосфотирозина с использованием антител. Рецепторные тирозинкиназы представляют собой важное семейство рецепторов клеточной поверхности, участвующих в передаче внеклеточных сигналов, таких как гормоны, факторы роста и цитокины. Связывание лиганда с тирозинкиназой мономерного рецептора стабилизирует взаимодействия между двумя мономерами с образованием димера, после чего два связанных рецептора фосфорилируют остатки тирозина в транс. Фосфорилирование и активация рецептора активирует сигнальный путь за счет ферментативной активности и взаимодействия с адапторными белками. Передача сигналов через рецептор эпидермального фактора роста (EGFR), рецепторную тирозинкиназу, имеет решающее значение для развития многих систем органов, включая кожу, легкие, сердце и мозг. Избыточная передача сигналов через путь EGFR обнаруживается при многих раковых заболеваниях человека.
Циклинзависимые киназы (CDK) представляют собой серин-треониновые киназы, которые регулируют прогрессирование через эукариотические клетки клеточный цикл. CDK каталитически активны только при связывании с регуляторным циклином. Клетки животных содержат по крайней мере девять различных CDK, которые со значительной специфичностью связываются с различными циклинами. Ингибиторы CDK (CKI) блокируют активность киназы в комплексе циклин-CDK, чтобы остановить клеточный цикл в G1 или в ответ на сигналы окружающей среды или повреждение ДНК. Активность различных CDK активирует клеточные сигнальные пути и факторы транскрипции, которые регулируют ключевые события митоза, такие как фазовый переход G1 / S. Более ранние комплексы циклин-CDK обеспечивают сигнал для активации последующих комплексов циклин-CDK.
В данной клетке существуют тысячи различных сайтов фосфорилирования, поскольку:
Поскольку фосфорилирование любого сайта в данном белке может изменить функцию или локализацию этого белка, для понимания «состояния» клетки необходимо знать фосфо состояние рилирования его белков. Например, если аминокислота серин-473 («S473») в белке AKT фосфорилирована, AKT, как правило, функционально активен как киназа. В противном случае это неактивная киназа.
Сайты фосфорилирования имеют решающее значение для белков, их транспортировки и функций. Они представляют собой ковалентную модификацию белков посредством обратимого фосфорилирования. Это позволяет белкам оставаться в клетке, поскольку отрицательный фосфорилированный сайт препятствует их проницаемости через клеточную мембрану. Дефосфорилирование белка позволяет клетке пополнять запасы фосфатов, высвобождение которых экономит использование АТФ в клетке. Пример фосфорилирующего фермента обнаружен в E. coli бактерии. Он содержит щелочную фосфатазу в области своей мембраны. Самая внешняя мембрана проницаема для фосфорилированных молекул, однако внутренняя цитоплазматическая мембрана непроницаема из-за больших отрицательных зарядов. Таким образом, бактерии E. coli накапливают белки и пирофосфаты в своей периплазматической мембране до тех пор, пока они не потребуются внутри клетки.
Недавние успехи в идентификации фосфопротеомиков привели к открытиям бесчисленных сайтов фосфорилирования в белках. Это потребовало интегративной среды для доступных данных, в которой организованы известные сайты фосфорилирования белков. Была создана тщательно подобранная база данных dbPAF, содержащая известные сайты фосфорилирования в H. sapiens, М. musculus, R. norvegicus, Д. melanogaster, C. elegans, S. pombe и S. cerevisiae. База данных в настоящее время содержит 294 370 неизбыточных сайтов фосфорилирования 40 432 белков. Другие инструменты прогнозирования фосфорилирования белков включают NetPhos для эукариот, NetPhosBac для бактерий и ViralPhos для вирусов.
Существует большое разнообразие остатков серина, и фосфорилирование каждого остатка может привести к различным метаболическим последствиям.
Фосфорилирование серина и Известно, что остатки треонина взаимодействуют с модификацией O-GlcNAc остатков серина и треонина.
Фосфорилирование тирозина быстро реагирует, и реакцию можно обратить. Являясь одним из основных в передаче сигнала - клеточном росте, дифференцировке, миграции, это клеточные процессы, поддерживаемые фосфорилированием тирозина. Функция протеинтирозинкиназ и протеин-тирозинфосфатазы уравновешивает уровень фосфотирозина на любом протеине. Нарушение работы определенных цепей протеинтирозинкиназ и протеинтирозинфосфатазы было связано с множеством заболеваний человека, такими как ожирение, инсулинорезистентность и сахарный диабет 2 типа. Фосфорилирование тирозина происходит не только у эукариот, но было обнаружено, что оно происходит у некоторых видов бактерий и присутствует среди прокариот. Фосфорилирование тирозина поддерживает клеточную регуляцию у бактерий, аналогичную его функции у эукариот.
Аргинин фосфорилирование во многих грамположительных бактериях маркирует белки для разложения с помощью Clp протеаза.
Недавние исследования лаборатории Claire E Eyers подтверждают широко распространенное фосфорилирование белков человека на множестве неканонических аминокислот, включая мотивы, содержащие фосфорилированный гистидин (1 и 3 положения), аспартат, цистеин, глутамат, аргинин и лизин в экстрактах клеток HeLa. Из-за химической и термической лабильности этих фосфорилированных остатков для сохранения наряду с термостабильным «классическим» фосфорилированием Ser, Thr и Tyr требуются специальные процедуры и методы разделения.
Антитела могут использоваться в качестве мощного инструмента для определения того, фосфорилируется ли белок в определенном месте. Антитела связываются с белком и обнаруживают вызванные фосфорилированием конформационные изменения. Такие антитела называются фосфоспецифическими антителами; сейчас доступны сотни таких антител. Они становятся важнейшими реагентами как для фундаментальных исследований, так и для клинической диагностики.
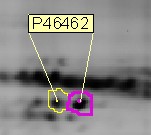 Пример посттрансляционной модификации, обнаруженной на 2D-геле (границы пятна ограничены программным обеспечением анализа, идентификация с помощью масс-спектрометрии, P46462 - идентификатор белка в Expasy)
Пример посттрансляционной модификации, обнаруженной на 2D-геле (границы пятна ограничены программным обеспечением анализа, идентификация с помощью масс-спектрометрии, P46462 - идентификатор белка в Expasy) Изоформы посттрансляционной модификации (PTM) легко обнаруживаются на 2D гели. Действительно, фосфорилирование заменяет нейтральные гидроксильные группы серинов, треонинов или тирозинов на отрицательно заряженные фосфаты с pKs около 1,2 и 6,5. Таким образом, при pH ниже 5,5 фосфаты добавляют один отрицательный заряд; около pH 6,5 они добавляют 1,5 отрицательных заряда; выше pH 7,5 они добавляют 2 отрицательных заряда. Относительное количество каждой изоформы также можно легко и быстро определить по интенсивности окрашивания на 2D-гелях.
В некоторых очень специфических случаях обнаружение фосфорилирования как сдвига в электрофоретической подвижности белка возможно на простых одномерных гелях SDS-PAGE, как это описано, например, для транскрипционного коактиватора Kovacs et al.. Считается, что сильные конформационные изменения, связанные с фосфорилированием (которые сохраняются в растворах, содержащих детергент), лежат в основе этого явления. Большинство сайтов фосфорилирования, для которых описан такой сдвиг подвижности, попадают в категорию сайтов SP и TP (т.е. остаток пролина следует за остатком фосфорилированного серина или треонина).
Совсем недавно крупномасштабные масс-спектрометрические анализы стали использовать для определения сайтов фосфорилирования белка. За последние 4 года были опубликованы десятки исследований, каждое из которых идентифицирует тысячи сайтов, многие из которых ранее не были описаны. Масс-спектрометрия идеально подходит для таких анализов с использованием HCD или ETD фрагментации, поскольку добавление фосфорилирования приводит к увеличению массы белка и фосфорилированного остатка. Для этих исследований необходимы современные высокоточные масс-спектрометры, поэтому технология ограничивается лабораториями с масс-спектрометрами высокого класса. Однако анализ фосфорилированных пептидов с помощью масс-спектрометрии все еще не так прост, как для «обычных» немодифицированных пептидов. Недавно был разработан EThcD, сочетающий перенос электрона и столкновительную диссоциацию при более высоких энергиях. По сравнению с обычными методами фрагментации, схема EThcD обеспечивает более информативные спектры МС / МС для однозначной локализации фосфозита.
Детальная характеристика сайтов фосфорилирования очень трудна, а количественное определение белка фосфорилирование с помощью масс-спектрометрии требует подходов изотопного внутреннего стандарта. Относительное количественное определение может быть получено с помощью различных технологий дифференциальной маркировки изотопов. Существует также несколько методов количественного фосфорилирования белков, включая флуоресцентные иммуноанализы, термофорез на микромасштабах, FRET, TRF, поляризацию флуоресценции, тушение флуоресценции, сдвиг подвижности, детекцию на основе гранул и клеточное определение. на основе форматов.
Фосфорилирование белков является обычным явлением для всех видов жизни, включая всех животных, растения, грибы, бактерии и археи. Происхождение механизмов фосфорилирования белков является наследственным и сильно различается между разными видами. У эукариот, по оценкам, от 30 до 65% всех белков могут быть фосфорилированы с десятками или даже сотнями тысяч отдельных участков фосфорилирования. Некоторые сайты фосфорилирования, по-видимому, эволюционировали как условные выключатели, блокирующие активный центр фермента, например, в прокариотическом метаболическом ферменте изоцитратдегидрогеназе. Однако в случае белков, которые должны быть фосфорилированы, чтобы быть активными, менее ясно, как они могли возникнуть от нефосфорилированных предков. Было показано, что подмножество сериновых фосфозитов часто заменяется кислотными остатками, такими как аспартат и глутамат, между различными видами. Эти анионные остатки могут взаимодействовать с катионными остатками, такими как лизин и аргинин, с образованием солевых мостиков, стабильных нековалентных взаимодействий, которые изменяют структуру белка. Эти фосфозиты часто участвуют в солевых мостиках, предполагая, что некоторые сайты фосфорилирования эволюционировали как условные переключатели для солевых мостиков, позволяя этим белкам принимать активную конформацию только в ответ на определенный сигнал.
Их ~ 600 известные эукариотические протеинкиназы, что делает их одним из крупнейших семейств генов. Большая часть фосфорилирования осуществляется одним суперсемейством протеинкиназ, которые разделяют консервативный домен киназы. Фосфорилирование белков является высококонсервативным в путях, имеющих ключевое значение для выживания клеток, таких как прогрессирование клеточного цикла, основанное на циклин-зависимых киназах (CDK), но отдельные сайты фосфорилирования часто бывают гибкими. Мишени фосфорилирования CDK часто имеют фосфозиты в неупорядоченных сегментах, которые находятся в неидентичных местах даже у близких видов. Напротив, мишени фосфорилирования CDK в структурно определенных областях более консервативны. Хотя активность CDK имеет решающее значение для роста и выживания клеток у всех эукариот, только очень немногие фосфозиты демонстрируют сильную консервацию их точного положения. Позиционирование, вероятно, будет очень важно для фосфатов, которые аллостерически регулируют структуру белка, но гораздо более гибким для фосфатов, которые взаимодействуют с фосфопептид-связывающими доменами для рекрутирования регуляторных белков.
Фосфорилирование белков - это обратимая посттрансляционная модификация белков. У эукариот фосфорилирование белков влияет на передачу сигналов, экспрессию генов и дифференцировку. Он также участвует в репликации ДНК во время клеточного цикла и в механизмах, которые справляются со стресс-индуцированными блоками репликации. По сравнению с эукариотами, прокариоты используют киназы и фосфатазы типа Хэнкса для передачи сигналов. Может ли фосфорилирование белков в бактериях также регулировать такие процессы, как репарация ДНК или репликация, все еще остается неясным.
По сравнению с фосфорилированием белков прокариот, исследования фосфорилирования белков у эукариот от дрожжей до клеток человека были скорее обширный. Известно, что эукариоты полагаются на фосфорилирование гидроксильной группы на боковых цепях серина, треонина и тирозина для передачи клеточных сигналов. Это основные регуляторные посттрансляционные модификации в эукариотических клетках, но фосфорилирование белков прокариот изучено менее интенсивно. В то время как серин, треонин и тирозин фосфорилируются у эукариот, гистидин и аспартат фосфорилируются у прокариот, растений и нерастительных эукариот. У бактерий фосфорилирование гистидина происходит в фосфоенолпируват-зависимых фосфотрансферазных системах (PTS), которые участвуют в процессе интернализации, а также в фосфорилировании сахаров.
Фосфорилирование белков протеинкиназой было впервые показано на E. coli и Salmonella typhimurium, но с тех пор было продемонстрировано на многих других бактериальных клетках. Было обнаружено, что бактерии используют фосфорилирование гистидина и аспартата в качестве модели для бактериальной сигнальной трансдукции, но в последние несколько лет появились доказательства того, что фосфорилирование серина, треонина и тирозина также присутствует в бактериях. Было показано, что бактерии несут киназы и фосфатазы, аналогичные их эукариотическим эквивалентам, но они также развили уникальные киназы и фосфатазы, которые не обнаруживаются у эукариот.
Аномальное фосфорилирование белков связано с ряд заболеваний, в частности рак, но также болезнь Альцгеймера, болезнь Паркинсона и другие дегенеративные заболевания.
тау-белок к группе белков, связанных с микротрубочками (MAP), которые, среди прочего, помогают стабилизировать микротрубочки в клетках, включая нейроны. Ассоциативная и стабилизирующая активность тау-белка зависит от его фосфорилированного состояния. При болезни Альцгеймера из-за неправильной укладки и аномальных конформационных изменений в структуре тау-белка он становится неэффективным при связывании с микротрубочками и, таким образом, неспособен поддерживать структуру нервного цитоскелета в организованном состоянии во время нервных процессов; фактически аномальный тау-белок ингибирует и нарушает организацию микротрубочек и отключает нормальный тау-белок от микротрубочек в цитозольную фазу. Неправильная укладка приводит к аномальной агрегации в фибриллярные клубки внутри нейронов, что является признаком болезни Альцгеймера. Существует достаточное количество, которое необходимо фосфорилированию тау-белка для функционирования, но гиперфосфорилирование тау-белка считается одним из основных факторов, влияющих на его неспособность связываться. Фосфатазы PP1, PP2A, PP2B и PP2C дефосфорилируют тау-белок in vitro, и их активность, как было обнаружено, снижена в областях мозга у пациентов с болезнью Альцгеймера. Фосфопротеин тау-белка гиперфосфорилируется в три-четыре раза у пациента с болезнью Альцгеймера по сравнению с пожилым человеком, не страдающим болезнью. Тау-белок болезни Альцгеймера, по-видимому, удаляет MAP1 и MAP2 (два других основных ассоциированных белка) из микротрубочек, и этот вредный эффект устраняется при дефосфорилировании, что свидетельствует о гиперфосфорилировании как единственной причине парализующей активности.
α-Синуклеин - это белок, связанный с болезнью Паркинсона. Этот белок кодируется геном PARRK1, и в своей нативной форме α-синуклеин участвует в рециркуляции синаптических пузырьков, которые несут нейротрансмиттеры, и в естественных условиях находится в развернутой форме. Повышенные уровни α-синуклеина обнаруживаются у пациентов с болезнью Паркинсона, и, по-видимому, существует положительная корреляция между количеством белка α-синуклеина, присутствующего у пациента, и тяжестью заболевания.
Фосфорилирование аминокислоты Ser в белке α-синуклеина оказывает сильное влияние на тяжесть заболевания. По-видимому, существует корреляция между общей концентрацией альфа-синуклеина (нефосфорилированного) и тяжестью симптомов у пациентов с болезнью Паркинсона. У здоровых пациентов уровень нефосфорилированного α-синуклеина выше, чем у пациентов с болезнью Паркинсона. Более того, измерение изменений соотношения концентраций фосфорилированного α-синуклеина к нефосфорилированному α-синуклеину у пациента может быть потенциальным маркером прогрессирования заболевания
Фосфорилирование Ser связано с агрегацией белок и дальнейшее повреждение нервной системы. Кроме того, агрегация фосфорилированного α-синуклеина может быть усилена, если пресинаптический каркасный белок Sept4 присутствует в недостаточных количествах. Важно отметить, что прямое взаимодействие α-синуклеина с белком Sept4 ингибирует фосфорилирование Ser.