
Войти
Фосфатидные кислоты - это анионные фосфолипиды, важные для передачи сигналов клеток и прямой активации липидно-зависимых ионных каналов. Гидролиз фосфатидной кислоты дает по одной молекуле глицерина и фосфорной кислоты и по две молекулы жирных кислот. Они составляют около 0,25% фосфолипидов в бислое.
 Общая химическая структура фосфатидных кислот
Общая химическая структура фосфатидных кислот Фосфатидная кислота состоит из глицериновой основной цепи с, как правило, насыщенной жирной кислотой, связанной с углеродом -1, ненасыщенная жирная кислота, связанная с углеродом -2, и фосфатная группа, связанная с углеродом -3.
Помимо синтеза de novo, PA может быть образован тремя способами:
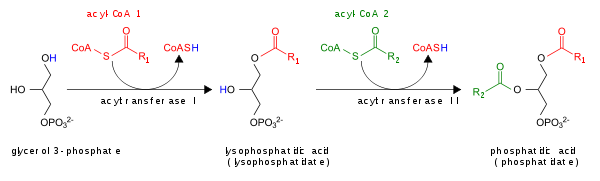
PA разрушается путем превращения в DAG с помощью липид-фосфатфосфогидролаз (LPP) или в лизо-PA с помощью фосфолипазы A (PLA).
Роль PA в клетке можно разделить на три категории:
Первые три роли не исключают друг друга. Например, PA может участвовать в образовании пузырьков, способствуя искривлению мембраны и рекрутируя белки для выполнения гораздо более энергетически невыгодной задачи образования шейки и защемления.
PA - это жизненно важный липид клетки, который действует как биосинтетический предшественник для образования (прямо или косвенно) всех липидов ацилглицерина в клетке.
In mam малийских и дрожжевых клеток известны два различных пути синтеза PA de novo: глицерин-3-фосфатный путь или дигидроксиацетонфосфатный путь. У бактерий присутствует только первый путь, и мутации, блокирующие этот путь, являются летальными, что демонстрирует важность PA. В клетках млекопитающих и дрожжей, где ферменты этих путей избыточны, мутация любого фермента не является летальной. Однако стоит отметить, что in vitro различные ацилтрансферазы проявляют различную субстратную специфичность в отношении ацил-КоА, которые включены в PA. Различные ацилтрансферазы также имеют различное внутриклеточное распределение, такое как эндоплазматический ретикулум (ER), митохондрии или пероксисомы, и локальные концентрации активированных жирных кислот. Это говорит о том, что различные ацилтрансферазы, присутствующие в клетках млекопитающих и дрожжей, могут быть ответственны за продуцирование разных пулов PA.
Превращение PA в диацилглицерин (DAG) с помощью LPP является этапом связывания для производство фосфатидилхолина (PC), фосфатидилэтаноламина (PE) и фосфатидилсерина (PS). Кроме того, DAG также превращается в CDP-DAG, который является предшественником фосфатидилглицерина (PG), фосфатидилинозитола (PI) и фосфоинозитидов (PIP, PIP 2, концентрации PIP 3).
PA поддерживаются на чрезвычайно низких уровнях в клетке за счет активности мощных LPP. Они очень быстро превращают PA в DAG, и, поскольку DAG является предшественником многих других липидов, он также вскоре метаболизируется в другие мембранные липиды. Это означает, что любое усиление продукции PA может быть согласовано со временем с соответствующей повышающей регуляцией LPP и ферментов, метаболизирующих DAG.
PA, следовательно, важен для синтез липидов и выживаемость клеток, тем не менее, в нормальных условиях поддерживаются на очень низком уровне в клетке.
PA - уникальный фосфолипид в том смысле, что он имеет небольшой высокий уровень заряженная головная группа, которая очень близка к глицериновой основной цепи. Известно, что PA играет роль как в делении и слиянии везикул , так и в слиянии везикул, и Эти роли могут относиться к биофизическим свойствам ПА.
В местах отрастания или слияния мембраны мембрана становится или сильно изогнута. Важным событием в отрастании пузырьков, таких как транспортные переносчики из Golgi, является создание и последующее сужение шейки мембраны. Исследования показали, что этот процесс может быть вызван липидами, и постулировали центральную роль DAG из-за его, также, уникальной молекулярной формы. Присутствие двух ацильных цепей, но отсутствие головной группы приводит к большой отрицательной кривизне мембран.
LPAAT BARS-50 также участвует в отпочковании от Golgi. Это говорит о том, что превращение лизоПА в PA может влиять на кривизну мембраны. Активность LPAAT удваивает количество ацильных цепей, значительно увеличивая площадь поперечного сечения липида, который находится «внутри» мембраны, в то время как поверхностная головная группа остается неизменной. Это может привести к более отрицательной кривизне мембраны. Исследователи из Утрехтского университета изучали влияние лизоРА по сравнению с PA на кривизну мембраны, измеряя их влияние на температуру перехода PE из липидных бислоев в неламеллярные фазы с помощью P-ЯМР. Было показано, что кривизна, индуцированная этими липидами, зависит не только от структуры лизоПА по сравнению с PA, но также от динамических свойств, таких как гидратация головных групп и меж- и внутримолекулярные взаимодействия. Например, Са может взаимодействовать с двумя ПА с образованием нейтрального, но сильно изогнутого комплекса. Нейтрализация в противном случае отталкивающих зарядов головных групп и отсутствие каких-либо стерических препятствий делает возможными сильные межмолекулярные взаимодействия между ацильными цепями, в результате чего образуются микродомены, богатые PA. Таким образом, in vitro физиологические изменения pH, температуры и концентраций катионов оказывают сильное влияние на кривизну мембраны, вызванную PA и lysoPA. Взаимное превращение лизоПА, PA и DAG - а также изменения pH и концентрации катионов - могут вызывать изгиб и дестабилизацию мембраны, играя прямую роль в делении мембран просто в силу своих биофизических свойств. Однако, хотя было показано, что PA и lysoPA влияют на кривизну мембраны in vitro; их роль in vivo неясна.
Роль lysoPA, PA и DAG в стимулировании кривизны мембраны не исключает их роли в рекрутировании белков на мембрану. Например, потребность в Са для слияния сложных липосом не сильно зависит от добавления аннексина I, хотя она снижается за счет PLD. Однако с аннексином I и PLD степень слияния значительно увеличивается, а потребность в Ca снижается почти в 1000 раз до почти физиологических уровней.
Таким образом, метаболическая, биофизическая, рекрутирующая и сигнальная роли PA могут быть взаимосвязаны.
Поскольку PA быстро преобразуется в DAG, он очень недолговечен в клетке. Это означает, что сложно измерить продукцию PA и, следовательно, изучить роль PA в клетке. Однако активность PLD можно измерить путем добавления в ячейку первичных спиртов. Затем PLD проводит реакцию трансфосфатидилирования вместо гидролиза с образованием фосфатидиловых спиртов вместо PA. Фосфатидиловые спирты являются тупиками метаболизма, и их можно легко экстрагировать и измерять. Таким образом, можно измерить активность PLD и продукцию PA (если не сам PA), и, блокируя образование PA, можно сделать вывод об участии PA в клеточных процессах.
PA сохраняется на низком уровне в объеме мембраны, чтобы временно взрываться и локально передавать сигнал в высокой концентрации. Например, каналы TREK-1 активируются локальной ассоциацией с PLD и производством PA. Константа диссоциации ПА для ТРЕК-1 составляет примерно 10 мкмоль. Относительно слабое связывание в сочетании с низкой концентрацией PA в мембране позволяет каналу отключаться. Высокая локальная концентрация для активации предполагает, по крайней мере, некоторые ограничения в локальной диффузии липидов. Основная низкая концентрация PA в сочетании с высокими локальными всплесками противоположна передаче сигналов PIP2. PIP2 удерживается на относительно высоком уровне в мембране и затем временно гидролизуется рядом с белком, чтобы временно снизить передачу сигналов PIP2. Передача сигналов PA отражает передачу сигналов PIP2 в том смысле, что основная концентрация сигнального липида не должна изменяться для оказания мощного местного эффекта на целевой белок.
Как описано выше, PLD гидролизует PC с образованием PA и холина. Поскольку холина в клетке очень много, активность PLD не оказывает значительного влияния на уровень холина; и холин вряд ли будет играть какую-либо роль в передаче сигналов.
Роль активации PLD во многих контекстах передачи сигналов, в сочетании с отсутствием роли холина, предполагает, что PA играет важную роль в передаче сигналов. Однако PA быстро превращается в DAG, и DAG также известен как сигнальная молекула. Это поднимает вопрос о том, играет ли PA какую-либо прямую роль в передаче сигналов или он просто действует как предшественник для продукции DAG. Если будет обнаружено, что PA действует только как предшественник DAG, тогда можно поднять вопрос, почему клетки должны производить DAG с использованием двух ферментов, если они содержат PLC, который может продуцировать DAG за один шаг.
PA, продуцируемый PLD или DAGK, можно отличить по добавлению [γ-P] ATP. Это покажет, является ли фосфатная группа новым производным от активности киназы или она происходит от ПК.
Хотя PA и DAG взаимопревращаемы, они не действуют одними и теми же путями. Стимулы, которые активируют PLD, не активируют ферменты ниже DAG, и наоборот. Например, было показано, что добавление PLD к мембранам приводит к продукции [P]-меченых PA и [P]-меченных фосфоинозитидов. Добавление ингибиторов DAGK устраняет продукцию [P]-меченого PA, но не стимулирует PLD-стимулированное производство фосфоинозитидов.
Возможно, что, хотя PA и DAG являются взаимопревращаемыми, отдельные пулы сигнальных и несигнальных липидов могут сохраняться. Исследования показали, что передача сигналов DAG опосредуется полиненасыщенным DAG, тогда как PA, происходящий из PLD, является мононенасыщенным или насыщенным. Таким образом, функциональный насыщенный / мононенасыщенный PA может быть разложен путем его гидролиза с образованием нефункционального насыщенного / мононенасыщенного DAG, тогда как функциональный полиненасыщенный DAG может быть разложен путем преобразования его в нефункциональный полиненасыщенный PA.
Эта модель предполагает, что PA и эффекторы DAG должны уметь различать липиды с одинаковыми головными группами, но с разными ацильными цепями. Хотя некоторые липид-связывающие белки способны внедряться в мембраны и гипотетически распознавать тип ацильной цепи или результирующие свойства мембраны, многие липид-связывающие белки являются цитозольными и локализуются на мембране, связывая только головные группы липидов. Возможно, различные ацильные цепи могут влиять на угол наклона головной группы в мембране. Если это так, это предполагает, что PA-связывающий домен должен быть не только способен специфически связывать PA, но также должен быть способен идентифицировать те головные группы, которые находятся под правильным углом. Каким бы ни был механизм, такая специфика возможна. Это наблюдается в DAGK семенников свиней, который специфичен для полиненасыщенного DAG, и в двух LPP гепатоцитов крысы, которые дефосфорилируют различные виды PA с различной активностью. Более того, было показано, что стимуляция активности SK1 с помощью PS in vitro сильно варьируется в зависимости от того, использовались ли диолеоил (C18: 1), дистеароил (C18: 0) или 1-стеароил, 2-олеоил разновидности PS. Таким образом, кажется, что, хотя PA и DAG взаимопревращаемы, разные виды липидов могут иметь различную биологическую активность; и это может позволить двум липидам поддерживать отдельные сигнальные пути.