
Войти
| Paramyxoviridae | |
|---|---|
 | |
| Организация вириона и генома вируса чумы собак (CDV) | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Царство: | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Негарнавирикота |
| Класс: | Monjiviricetes |
| Заказ: | Mononegavirales |
| Семья: | Paramyxoviridae |
| Подсемейства | |
| |
Paramyxoviridae (от греч. Пара- «рядом с» и myxa « слизь ») представляет собой семейство РНК-вирусов с отрицательной цепью в отряде Mononegavirales. Позвоночные животные служат естественными хозяевами. Заболевания, связанные с этим семейством, включают корь, паротит и инфекции дыхательных путей. В семействе четыре подсемейства, 17 родов и 78 видов, три рода из которых не относятся к подсемейству.
Вирионы имеют оболочку, могут быть сферическими или плеоморфными и способны продуцировать нитчатые вирионы. Диаметр около 150 нм. Геномы линейны, длиной около 15 килобайт. Слитые белки и белки прикрепления появляются в виде шипов на поверхности вириона. Белки матрицы внутри оболочки стабилизируют структуру вируса. Ядро нуклеокапсида состоит из геномной РНК, белков нуклеокапсида, фосфопротеинов и белков полимеразы.
 Структура генома парамиксовируса
Структура генома парамиксовируса Геном является нерасчлененными, отрицательно-смысловой РНК, 15-19 т.п.н. в длину и содержит от шести до 10 генов. Экстрацистронные (некодирующие) области включают:
Каждый ген содержит сигналы начала / остановки транскрипции в начале и в конце, которые транскрибируются как часть гена.
Последовательность генов внутри генома сохраняется во всем семействе из-за явления, известного как полярность транскрипции (см. Mononegavirales ), при котором гены, расположенные ближе всего к 3'-концу генома, транскрибируются в большем количестве, чем гены ближе к 5'-концу. Это результат структуры генома. После того, как каждый ген транскрибируется, РНК-зависимая РНК-полимераза делает паузу, чтобы высвободить новую мРНК, когда она встречает межгенную последовательность. Когда полимераза РНК приостановлена, существует шанс, что она отделится от генома РНК. Если он диссоциирует, он должен повторно войти в геном в лидерной последовательности, а не продолжать транскрибировать длину генома. В результате чем дальше от лидерной последовательности находятся гены, расположенные ниже по течению, тем меньше они будут транскрибироваться РНК-полимеразой.
Доказательства для модели единственного промотора были проверены, когда вирусы подвергались воздействию ультрафиолетового света. УФ-излучение может вызвать димеризацию РНК, что препятствует транскрипции РНК-полимеразой. Если вирусный геном следует модели с множественными промоторами, уровень ингибирования транскрипции должен коррелировать с длиной гена РНК. Однако лучше всего геном описывался с помощью модели с одним промотором. Когда геном парамиксовируса подвергался воздействию УФ-света, уровень ингибирования транскрипции был пропорционален расстоянию от лидерной последовательности. То есть, чем дальше ген от лидерной последовательности, тем больше вероятность того, что димеризация РНК ингибирует РНК-полимеразу.
Вирус использует модель с одним промотором, поскольку его гены расположены в относительном порядке белков, необходимом для успешного заражения. Например, нуклеокапсидный белок N необходим в большем количестве, чем РНК-полимераза L.
Вирусы семейства Paramyxoviridae также являются антигенно стабильными, что означает, что гликопротеины вирусов согласованы между различными штаммами одного и того же типа. Выдвигаются две причины этого явления: первая заключается в том, что геном не сегментирован, поэтому не может подвергнуться генетической реассортации. Чтобы этот процесс происходил, сегменты, необходимые для повторной сортировки, происходят, когда сегменты из разных штаммов смешиваются вместе, чтобы создать новый штамм. Без сегментов ничего нельзя смешивать друг с другом, поэтому антигенный сдвиг не происходит. Вторая причина связана с идеей антигенного дрейфа. Поскольку РНК-зависимая РНК-полимераза не имеет функции проверки ошибок, многие мутации происходят при процессинге РНК. Эти мутации накапливаются, и в конечном итоге создаются новые штаммы. Исходя из этой концепции, можно было бы ожидать, что парамиксовирусы не должны быть антигенно стабильными; однако, как представляется, верно обратное. Основная гипотеза, лежащая в основе антигенной стабильности вирусов, заключается в том, что каждый белок и аминокислота выполняют важную функцию. Таким образом, любая мутация приведет к снижению или полной потере функции, что, в свою очередь, приведет к снижению эффективности нового вируса. Эти вирусы не смогут выжить так долго, как более вирулентные штаммы, и поэтому вымрут.
Многие геномы парамиксовирусов следуют «правилу шести». Общая длина генома почти всегда кратна шести. Вероятно, это связано с преимуществом связывания всей РНК с белком N (поскольку N связывает гексамеры РНК). Если оставить РНК открытой, вирус не будет эффективно реплицироваться. Последовательность гена:
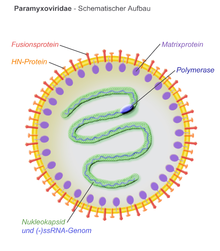 Иллюстрация вириона Paramyxoviridae
Иллюстрация вириона Paramyxoviridae  Репликация цикла вируса чумы собак (CDV).
Репликация цикла вируса чумы собак (CDV). Репликация вируса происходит в цитоплазме. Попадание в клетку-хозяин достигается за счет прикрепления вируса к клетке-хозяину. Репликация следует модели репликации вируса с отрицательной цепью РНК. Транскрипция вируса с отрицательной цепью РНК с использованием полимеразного заикания является методом транскрипции. Трансляция происходит посредством сканирования с утечкой информации, рибосомного шунтирования и повторной инициации терминации РНК. Вирус выходит из клетки-хозяина почкованием. Естественными хозяевами служат люди, позвоночные и птицы. Путь передачи - частицы в воздухе.
Paramyxoviridae способны пройти редактирование мРНК, которая производит различные белки из того же транскрипта мРНК путем смещать назад одно основание, чтобы прочитать в другом открытой рамки считывания ( ORF ) в связи с наличием вторичных структур, таких как псевдоузлы. Paramyxoviridae также претерпевают заикание трансляции для образования поли (А) хвоста на конце транскриптов мРНК, многократно перемещаясь назад по одному нуклеотиду за раз в конце матрицы РНК.
 Филогенетическое дерево парамиксовирусов
Филогенетическое дерево парамиксовирусов Семья: Paramyxoviridae
Парамиксовирусы вызывают ряд серьезных заболеваний человека. К ним относятся эпидемический паротит, а также корь, от которых в 2000 году умерло около 733 000 человек.
В вирусы парагриппа человека (HPIV) являются второй наиболее частой причиной заболевания дыхательных путей у младенцев и детей. Существует четыре типа HPIV, известных как HPIV-1, HPIV-2, HPIV-3 и HPIV-4. HPIV-1 и HPIV-2 могут вызывать симптомы простуды, а также круп у детей. HPIV-3 связан с бронхиолитом, бронхитом и пневмонией. HPIV-4 встречается реже, чем другие типы, и, как известно, вызывает легкие и тяжелые заболевания дыхательных путей.
Парамиксовирусы также несут ответственность за ряд заболеваний у других видов животных, например вирус чумы собак ( собаки ), вирус чумы фосиновых ( тюлени ), морбилливирус китообразных ( дельфины и морские свиньи ), вирус болезни Ньюкасла ( птицы ) и вирус чумы крупного рогатого скота ( крупный рогатый скот). ). Некоторые парамиксовирусы, такие как генипавирусы, являются зоонозными патогенами, встречающимися в природе в организме животного-хозяина, но также способными инфицировать людей.
Вирус Hendra (HeV) и вирус Nipah (NiV) из рода Henipavirus появились у людей и домашнего скота в Австралии и Юго-Восточной Азии. Оба вируса заразны, очень вирулентны и способны инфицировать ряд видов млекопитающих и вызывать потенциально смертельные заболевания. Из-за отсутствия лицензированной вакцины или противовирусной терапии HeV и NiV обозначены как агенты уровня биобезопасности (BSL) 4. Геномная структура обоих вирусов аналогична типичной парамиксовирусу.
За последние несколько десятилетий парамиксовирусы были обнаружены у наземных, летучих и водных животных, продемонстрировав широкий диапазон хозяев и большое генетическое разнообразие вирусов. По мере развития молекулярных технологий и внедрения программ эпиднадзора за вирусами обнаружение новых вирусов в этой группе увеличивается.
Эволюция парамиксовирусов все еще обсуждается. Используя пневмовирусы (мононегавирусное семейство Pneumoviridae ) в качестве внешней группы, парамиксовирусы можно разделить на две клады: одну, состоящую из авулавирусов и рубулавирусов, и другую, состоящую из респиравирусов, генипавирусов и морбилливирусов. Во второй кладе респираторовирусы представляют собой базальную группу. Клада респиравирус-генпавирус-морбилливирус может быть базальной кладой авулавируса-рубулавируса.