
Войти
 | |
 | |
| Имена | |
|---|---|
| Предпочтительное название IUPAC Оксобутандиовая кислота | |
| Другие имена Щавелевоуксусная кислота Щавелевоуксусная кислота 2-Оксоянтарная кислота Кетоянтарная кислота | |
| Идентификаторы | |
| Количество CAS | |
| 3D модель ( JSmol ) | |
| ЧЭБИ | |
| ChemSpider | |
| ECHA InfoCard | 100.005.755 |
| Номер ЕС | |
| IUPHAR / BPS | |
| PubChem CID | |
| UNII | |
| Панель управления CompTox ( EPA) | |
ИнЧИ
| |
Улыбки
| |
| Характеристики | |
| Химическая формула | С 4 Н 4 О 5 |
| Молярная масса | 132,07 г / моль |
| Плотность | 0,18 г / см 3 |
| Температура плавления | 161 ° С (322 ° F, 434 К) |
| Термохимия | |
| Std энтальпия формации (Δ F H ⦵298) | -943,21 кДж / моль |
| Std энтальпии сгорания (Δ с Н ⦵298) | -1205,58 кДж / моль |
| Если не указано иное, данные приведены для материалов в их стандартном состоянии (при 25 ° C [77 ° F], 100 кПа). | |
| | |
| Ссылки на инфобоксы | |
Щавелевоуксусная кислота (также известная как щавелевоуксусная кислота или OAA) представляет собой кристаллическое органическое соединение с химической формулой HO 2 CC (O) СН 2 СО 2 Н. щавелевоуксусной кислота, в форме его сопряженного основание оксалоацетата, является метаболической промежуточной во многом процессы, происходящие у животных. Он принимает участие в глюконеогенезе, то цикл мочевины, то глиоксилатный цикл, аминокислоты, синтез, жирные кислоты, синтез и кислота цикл лимонного.
Щавелевоуксусная кислота подвергается последовательному депротонированию с образованием дианиона:
При высоком pH енолизируемый протон ионизируется:
В енольных формах щавелевоуксусной кислоты являются особенно стабильными, так что эти два таутомеры имеют разные точки плавления (152 ° С для цис изоформы и 184 ° C для транса - изоформов). Эта реакция катализируется ферментом оксалоацетаттаутомеразой. транс- энол-оксалоацетат также появляется, когда тартрат является субстратом для фумаразы.
 Оксалоацетаттаутомераза катализирует образование енолоксалоацетата. показана транс- изоформа.
Оксалоацетаттаутомераза катализирует образование енолоксалоацетата. показана транс- изоформа. Оксалоацетат в природе образуется несколькими способами. Основной маршрут при окислении из L -malate, катализируется малатдегидрогеназой, в цикле лимонной кислоты. Малат также окисляется сукцинатдегидрогеназой в медленной реакции, причем исходным продуктом является енолоксалоацетат. Он также возникает в результате конденсации пирувата с угольной кислотой, вызванной гидролизом АТФ :
Происходя в мезофилле растений, этот процесс протекает через фосфоенолпируват, катализируемый фосфоенолпируваткарбоксилазой. Оксалоацетат также может образовываться в результате транс- или деаминирования аспарагиновой кислоты.
Оксалоацетат является промежуточным продуктом цикла лимонной кислоты, где он реагирует с ацетил-КоА с образованием цитрата, катализируемого цитрат-синтазой. Он также участвует в глюконеогенезе, в цикле мочевины, то цикл глиоксилата, аминокислоты синтеза и жирные кислоты синтез. Оксалоацетат также является мощным ингибитором комплекса II.
Глюконеогенез - это метаболический путь, состоящий из серии из одиннадцати катализируемых ферментами реакций, приводящих к образованию глюкозы из неуглеводных субстратов. Начало этого процесса происходит в матриксе митохондрий, где находятся молекулы пирувата. Молекула пирувата карбоксилируется ферментом пируваткарбоксилазы, активируемым каждой молекулой из АТФ и воды. Эта реакция приводит к образованию оксалоацетата. НАДН восстанавливает оксалоацетат до малата. Это преобразование необходимо для транспортировки молекулы из митохондрий. Попав в цитозоль, малат снова окисляется до оксалоацетата с помощью НАД +. Тогда оксалоацетат остается в цитозоле, где и будут проходить остальные реакции. Оксалоацетат позже декарбоксилируется и фосфорилируется фосфоенолпируваткарбоксикиназой и становится 2-фосфоенолпируватом с использованием гуанозинтрифосфата (ГТФ) в качестве источника фосфата. Глюкоза получается после дальнейшей переработки.
Цикл мочевины - это метаболический путь, который приводит к образованию мочевины с использованием одной молекулы аммония из разложенных аминокислот, другой группы аммония из аспартата и одной молекулы бикарбоната. Этот путь обычно происходит в гепатоцитах. Реакции, связанные с циклом мочевины, производят НАДН, и НАДН можно получить двумя разными способами. В одном из них используется оксалоацетат. В цитозоле находятся молекулы фумарата. Фумарат может быть преобразован в малат под действием фермента фумаразы. Малат подвергается действию малатдегидрогеназы, превращаясь в оксалоацетат с образованием молекулы НАДН. После этого оксалоацетат будет переработан в аспартат, поскольку трансаминазы предпочитают эти кетокислоты другим. Эта рециркуляция поддерживает приток азота в ячейку.
 Связь щавелевоуксусной кислоты, яблочной кислоты и аспарагиновой кислоты
Связь щавелевоуксусной кислоты, яблочной кислоты и аспарагиновой кислоты Цикл глиоксилата представляет собой вариант кислотного цикла лимонной. Это анаболический путь, происходящий у растений и бактерий с использованием ферментов изоцитратлиаза и малатсинтаза. Некоторые промежуточные этапы цикла немного отличаются от цикла лимонной кислоты; тем не менее оксалоацетат выполняет ту же функцию в обоих процессах. Это означает, что оксалоацетат в этом цикле также действует как первичный реагент и конечный продукт. Фактически оксалоацетат является чистым продуктом глиоксилатного цикла, потому что его петля цикла включает две молекулы ацетил-КоА.
На предыдущих стадиях ацетил-КоА переносится из митохондрий в цитоплазму, где находится синтаза жирных кислот. Ацетил-КоА транспортируется в виде цитрата, который ранее был образован в матриксе митохондрий из ацетил-КоА и оксалоацетата. Эта реакция обычно инициирует цикл лимонной кислоты, но когда нет потребности в энергии, она транспортируется в цитоплазму, где расщепляется на цитоплазматический ацетил-КоА и оксалоацетат.
Другая часть цикла требует НАДФН для синтеза жирных кислот. Часть этой восстанавливающей способности генерируется, когда цитозольный оксалоацетат возвращается в митохондрии, пока внутренний митохондриальный слой не проницаем для оксалоацетата. Сначала оксалоацетат восстанавливается до малата с помощью НАДН. Затем малат декарбоксилируется до пирувата. Теперь этот пируват может легко попасть в митохондрии, где он снова карбоксилируется до оксалоацетата под действием пируваткарбоксилазы. Таким образом, перенос ацетил-КоА из митохондрий в цитоплазму приводит к образованию молекулы НАДН. Общую реакцию, которая носит спонтанный характер, можно резюмировать следующим образом:
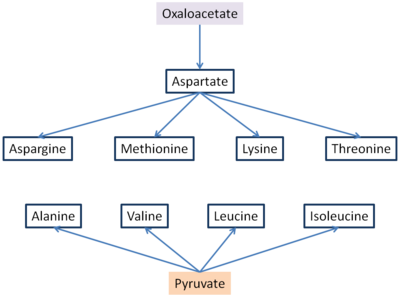
Шесть незаменимых аминокислот и три заменимых синтезируются из оксалоацетата и пирувата. Аспартат и аланин образуются из оксалоацетата и пирувата, соответственно, путем трансаминирования из глутамата. Аспарагин, метионин, лизин и треонин синтезируются аспартатом, поэтому, учитывая важность оксалоацетата, без него не будет образовываться аспартат и не будут образовываться следующие другие аминокислоты.
Оксалоацетат производит оксалат путем гидролиза.
Этот процесс катализируется ферментом оксалоацетазой. Этот фермент присутствует в растениях, но неизвестен в животном мире.
Нажмите на гены, белки и метаболиты ниже, чтобы ссылки на соответствующие статьи. [[Файл:  [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] | alt = Гликолиз и глюконеогенез править ]] Гликолиз и глюконеогенез править [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] | alt = Гликолиз и глюконеогенез править ]] Гликолиз и глюконеогенез править | Нажмите на гены, белки и метаболиты ниже, чтобы ссылки на соответствующие статьи. [[Файл:  [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] | alt = TCACycle_WP78 править ]] TCACycle_WP78 править [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] [[]] | alt = TCACycle_WP78 править ]] TCACycle_WP78 править |
| |||
|---|---|---|---|
| |      | ||
| | | НАДН + Н + + СО 2 | |
| CoA | НАД + | ||
 ЧАС 2О ЧАС 2О  цис- Аконитат ЧАС 2О НАД (Ф) + НАД (Ф) Н + Н + НАД (Ф) + НАД (Ф) Н + Н +  CO 2 CO 2  | |||