
Войти
| Ornitholestes. Временной диапазон: Поздний юрский период, 154 Ma Pre11 Ꞓ O S D C P T J K Pg N ↓ | |
|---|---|
 | |
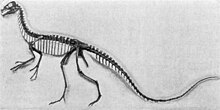
| Скелет Маунт, Королевский музей Тиррелла | |
| Научная классификация | |
| Царство: | Animalia |
| Тип: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Clade: | Maniraptoromorpha |
| Подсемейство: | †Ornitholestinae. Paul, 1988 |
| Род: | †Ornitholestes. Osborn, 1903 |
| Типовой вид | |
| †Ornitholestes hermanni . Осборн, 1903 | |
Ornitholestes (что означает «птичий грабитель») - небольшой теропод динозавр поздней юры (пачка Brushy Basin из формации Моррисон, средний кимериджский возраст, около 154 миллионов лет назад) западной Лавразия (территория, которая должна была стать Северной Америкой ).
На сегодняшний день Орнитолестес известен только по одному частичному скелету с сильно раздробленным черепом, найденным в каменоломне Костяной хижины рядом с Медисин-Боу, Вайоминг в 1900 году. Он был описан Генри Фэйрфилд Осборном в 1903 году. Незаконченная рука была позже приписана Орнитолесту, хотя теперь, похоже, принадлежит Таниколагреусу. Тип (и единственный известный) вид - это O. hermanni. Особое имя удостоено имени Американского музея естественной истории, подготовившего Адама Херманна.
 Восстановление жизни орнитолеста
Восстановление жизни орнитолеста Орнитолестес был двуногим хищником. Его голова была пропорционально меньше, чем у большинства других хищных динозавров, но череп был плотно сложен, с короткой мордой и крепкой нижней челюстью. орбиты (глазницы) были довольно большими, составляя более 25 процентов длины черепа. Нет никаких признаков костного кольца на глазу.
. Передние зубы Ornitholestes были несколько коническими, с уменьшенными зазубринами; задние зубы были загнутыми и более острыми, как у других динозавров-теропод. Генри Фэрфилд Осборн (1903) насчитал четыре зуба в предчелюстной кости, из которых передний зуб был самым большим в верхней челюсти. Напротив, Грегори С. Пол (1988) изобразил череп только с тремя оставшимися предчелюстными зубами, намного меньшими, чем те, что проиллюстрированы Осборном. Каждая верхнечелюстная кость (основная зубная кость верхней челюсти) содержала десять зубов, а каждая зубная кость (зубная кость нижней челюсти) содержала двенадцать зубов. Зубные ряды Ornitholestes были короткими, при этом зубной (нижний) ряд был даже короче, чем верхний (верхний) ряд, хотя сама зубная кость была исключительно длинной сзади, достигая точки ниже середины глазницы. Зубы не доходили до орбит, и ни один из зубных рядов не занимал более одной трети черепа.
 Иллюстрация голотипа черепа
Иллюстрация голотипа черепа Появляется область сломанной кости около наружной ноздри (ноздри) выпуклость вверх, что побудило Грегори С. Пола предположить в «Хищных динозаврах мира» (1988), что у орнитолеста был носовой рог, «похожий по внешнему виду на куриный гребень». Оба Oliver W.M. Rauhut (2003) и Kenneth Carpenter et al. (2005a) отвергли эту интерпретацию и указали, что вздутие кости вверх произошло из-за посмертного раздробления черепа. Обновленная иллюстрация Орнитолестеса Полом в его Полевом Путеводителе по динозаврам в Принстоне 2010 года больше не содержит носового рога.
Орнитолестес имел относительно короткую шею с небольшой сигмовидной (S-образной) кривой. Хвост был длинным и напоминающим хлыст, составлял более половины длины тела. Не все позвонки сохранились, но Осборн (1917) подсчитал, что у Ornitholestes было девять или десять шейных (шейных) позвонков, тринадцать спинных (спинных) позвонков, четыре крестцовых (бедренных) позвонка и от 39 до 44 хвостовых (хвостовых) позвонков. Карпентер и др. (2005a) зафиксировали, что образец содержал пять крестцовых позвонков. Ornitholestes был тероподом с коротким телом, и это отражалось в коротких размерах шейных и спинных позвонков спереди назад.
 Рука голотипа с восстановленным первым пальцем справа
Рука голотипа с восстановленным первым пальцем справа Передние конечности Орнитолесты были относительно длинными, чуть менее двух третей длины задних ног. плечевая кость (кость плеча) была сильно сложена и несколько длиннее, чем radius и ulna (кости предплечья). И плечевая кость, и лучевая кость имели прямой стержень. Когти на I и II пальцах руки были примерно одинакового размера. Хотя третья ногтевая кость руки (когтистая кость) не сохранилась, экстраполяция ближайших родственников Ornitholestes показывает, что она, вероятно, была короче первых двух.
Ornitholestes часто изображается как быстрый длинноногий теропод, но кости его нижних конечностей были довольно короткими. Осборн (1917) подсчитал, что отсутствующая большеберцовая кость (большеберцовая кость) составляла только около 70,6% длины бедренной кости (бедренной кости). плюсневые кости (кости стопы) расположены близко друг к другу, но не срослись. Как это типично для теропод, лапы были трехпалые (с тремя когтистыми несущими пальцами). Джон Х. Остром (1969) отметил, что коготь II пальца (самый внутренний палец) был больше, чем коготь III и IV, и предположил, что на этом пальце мог быть видоизмененный серповидный коготь, подобный когтю Дейнониха. Однако, как отметили и Остром (1969), и Пол (1988), плохая сохранность пальца II затрудняет подтверждение этой гипотезы.
 Сравнение размеров Ornitholestes
Сравнение размеров Ornitholestes В своем описании 1903 года Осборн писал, что длина Ornitholestes вдоль «черепа и позвоночника в восстановленном виде» составлял 2,22 м (7,28 фута). Однако эта реконструкция была неточной и частично основана на восстановлении Отниэлем Чарльзом Маршем базального зауроподоморфа Anchisaurus и шея и туловище были слишком удлиненными. Дэвид Норман (1985) и Джон Фостер (2007) подсчитали, что Орнитолестес был около 2 м в длину. В книге Грегори С. Пола «Хищные динозавры мира 1988 года» длина орнитолестов была указана примерно как 2,08 м (6,82 фута).
Пол (1988) и Фостер (2007) подсчитали, что Орнитолестес весил 12,6 кг (27,8 фунта).. Джон А. Лонг и Питер Схоутен (2008) предложили немного более высокую цифру, 15 кг (33 фунта).
 голотипу
голотипу Орнитолестес было первый теропод, обнаруженный в 1900-х годах. голотип скелет (AMNH 619) был обнаружен в июле 1900 г. в каменоломне Bone Cabin в Вайоминг Американским музеем естественной истории экспедиция Питера К. Кайзена, Пола Миллера и Фредерика Брюстера Лумиса. Он представляет собой частичный скелет с черепом, включающим многочисленные элементы позвоночника, передних конечностей, таза и задних конечностей. Генри Фэйрфилд Осборн назвал и научно описал этот экземпляр в 1903 году. Название рода Ornitholestes, первоначально предложенное Теодором Гиллом, означает «птичий грабитель» и происходит от греческого ὄρνις / ornis, ornithos («птица») и λῃστήσ / lestes («грабитель»). Название вида (O. hermanni) дано в честь Адама Германа, главного подготовителя в Музее, руководившего реставрацией и установкой скелета.
Была присвоена неполная рука (AMNH 587) to Ornitholestes Осборном в его описании рода 1903 г. Однако, как заметил Грегори С. Пол (Gregory S. Paul, 1988), плохая сохранность соответствующих элементов в типовом образце сделала эту ассоциацию «предварительной». В 2005 году Кеннет Карпентер и др. описал нового маленького теропода, Tanycolagreus, чей скелет был найден в карьере Костяной хижины всего в нескольких сотнях ярдов от AMNH 587. Поскольку AMNH 587 был практически идентичен сохранившейся руке экземпляра типа Tanycolagreus, теперь он является считается принадлежащим этому динозавру, а не орнитолестам. После этого переназначения Фил Сентер (2006) отметил, что «наши знания об Ornitholestes теперь можно почерпнуть только из голотипа». Джон Фостер (2007) сообщил, что некоторые фрагменты из карьера Dry Mesa могут принадлежать орнитолесту, хотя они еще не описаны.
В 1920 Чарльз Уитни Гилмор пришел к выводу, что Ornitholestes был идентичен Coelurus ; в 1934 году Оливер Перри Хэй признал разницу только на уровне видов, назвав Coelurus hermanni, но в 1980 году Джон Остром возродил этот род.
инфраотряд целурозаврия, изобретенный в 1914 году Фридрихом фон Хуэн, традиционно был таксономической корзиной для мусора, в которую помещались все мелкие тероподы. размещен. Таким образом, Ornitholestes из-за своего небольшого размера обычно классифицируется как целурозавр. В 1986 г. Жак Готье дал новое определение этому и нескольким другим палеонтологическим терминам более строго, основываясь на кладистических методах. Tetanurae был определен как современные птицы, и все тероподы более тесно связаны с современными птицами, чем с цератозаврами, в то время как целурозавры теперь включали всех представителей Tetanurae, более близких к современным птицам. птиц, чем карнозавров. В 1988 году Грегори С. Пол предположил, что орнитолестес очень похож по строению черепа на Proceratosaurus, теропод из средней юры из Англии. Он поместил эти два рода вместе в Ornitholestinae - новое подсемейство под Allosauridae - и предположил, что они более тесно связаны с гораздо более крупными Allosaurus, чем с другими маленькими тероподами. Однако классификация орнитолестов и процератозавров как родственников аллозавров оказалась несостоятельной (последний с тех пор оказался тираннозавроидом ), и Павел в конце концов отказался от нее. Все опубликованные кладистические анализы показали, что орнитолестес является целурозавром по определению Готье. Некоторый анализ подтвердил гипотезу о том, что это самый примитивный член группы Maniraptora, хотя более тщательный анализ показал, что он более примитивен, чем Maniraptoriformes, и, возможно, близок к нему. родственник "compsognathid" Juravenator starkii.
 Крепление голотипа скелета в начале 20 века на AMNH, которое почти не изменилось и сегодня
Крепление голотипа скелета в начале 20 века на AMNH, которое почти не изменилось и сегодня Следующее генеалогическое древо иллюстрирует синтез скелета взаимоотношения основных групп целурозавров, основанные на различных исследованиях, проведенных в 2010-х гг.
| Целурозаврия |
| ||||||||||||||||||||||||||||||||||||||||||||||||
В исследовании 2001 года, проведенном Брюсом Ротшильдом и другими палеонтологами, были исследованы 20 костей стопы, относящиеся к Ornitholestes стресс-разрушение, но не e были обнаружены.
Биомеханическое исследование, проведенное Филом Сентером (2006), использовало шарнирные слепки правой передней конечности образца типа Ornitholestes для определения диапазона его движений. Сентер обнаружил, что переднее плечо (предплечье) может свободно качаться в диапазоне 95 °. Когда согнуты (согнуты внутрь) в локтевом суставе до максимально возможной степени, плечевая кость (кость плеча) и лучевая кость (кость нижней руки) образуют угол 53 °. Способность Ornitholestes сгибать предплечье под углом, значительно более острым, чем 90 °, характерна для Maniraptoriformes, но отсутствует у более примитивных теропод, таких как Coelophysis и Allosaurus.
Даже когда полностью вытянутый (выпрямленный) в локте, предплечье не образует прямого угла, меньше этого угла на 22 °. Пронация (скручивание в ладонную сторону руки вниз) предплечья было невозможно, потому что лучевая и локтевая области не имели поверхностей качения, а это означало, что предплечье было в постоянном состоянии супинации.
Когда Орнитолестес сгибал локти, это заставляло предплечья двигаться внутрь, к средней линии. Возможно, он использовал эту способность, чтобы схватить добычу обеими руками одновременно.
 На этой иллюстрации Чарльза Р. Найта изображены охотящиеся на орнитолеста археоптериксы, хотя они не современники
На этой иллюстрации Чарльза Р. Найта изображены охотящиеся на орнитолеста археоптериксы, хотя они не современники Генри Фэйрфилд Осборн в своем описании орнитолеста в 1903 году отметил его большие конические передние зубы, «быструю хватательную силу» его руки и «уравновешивающую силу» его хвоста; он интерпретировал это как приспособление к добыче современных птиц. Позже Осборн опроверг эту гипотезу, предположив в 1917 году, что Орнитолестес представлял ранние стадии перехода от плотоядного к более травоядному образу жизни, но не раньше Чарльз Р. Найт нарисовал влиятельную и широко опубликованную иллюстрацию Орнитолестеса. в погоне за археоптериксом. Иллюстрация Найта и другие, заимствованные из нее, продолжали появляться в книгах о динозаврах, нацеленных на широкую публику на протяжении всего 20 века.
Дэвид Норман (1985) признал, что «вполне возможно, хотя и маловероятно», что Орнитолестес могли поймать и съесть примитивных птиц. Однако самые последние авторы вместо этого предложили диету из мелких наземных позвоночных. Млекопитающие, ящерицы, лягушки, саламандры, ринхоцефалы и только что вылупившиеся динозавры могли стать потенциальной добычей. Грегори С. Пол (1988) полагал, что Ornitholestes мог использовать свои конические передние зубы для ловли рыбы. Норман (1985) предположил, что крепкий череп и челюсти могли позволить орнитолестам схватить «более крупную и более активную добычу», чем другие маленькие теропод. В этом ключе Дэвид Ламберт (1993) предположил, что орнитолестес, если бы он был стайным охотником, мог бы «одолеть орнитопод размером с полувзрослый камптозавр."
в В 2007 году Джон Фостер, специалист по формации Моррисон, предположил возможность разделения ниши между Ornitholestes и его современным Coelurus, который был в пределах того же диапазона размеров. Он предположил, что большеглазые Ornitholestes могли быть специализированными для ночной охоты, тогда как Coelurus, возможно, сосредоточился на тех видах добычи, которые были активны в течение дня. Фостер отметил, однако, что эта гипотеза была в значительной степени спекулятивной; отсутствие сохранившегося материала черепа от Coelurus делает невозможным проверьте, были ли его глаза пропорционально меньше, чем у орнитолестов.
Пол (1988) подсчитал, что у 12-килограммовых (26 фунтов) эндотермических орнитолестов суточная потребность в рационе составляет около 700 г ( 1,54 фунта) плоти.
Орнит гололог Перси Лоу предположил в 1944 году, что Орнитолестес мог иметь перья. Однако эта интерпретация в основном игнорировалась более полувека; большинство реконструкций динозавров-теропод, включая Ornitholestes, изображали их с чешуйчатой кожей рептилий. Одним из немногих исключений из этого правила был труд Грегори С. Пола «Хищные динозавры мира» (1988). Роберт Т. Баккер «Ереси динозавров» (1986) точно предсказал присутствие перья на дромеозавридах и утверждал, что все динозавры были эндотермическими, но не использовал перья в иллюстрации Ornitholestes.
В 1996 году в Китае был обнаружен примитивный целурозавр Sinosauropteryx. Хорошо сохранившаяся окаменелость показала, что этот динозавр был покрыт рудиментарными меховыми перьями. В результате этого и других подобных открытий большинство палеонтологов теперь считают вероятным, что все целурозавры имели те или иные изолирующие перья или произошли от предков, которые имели их. Основываясь на его филогенетическом положении, Джон Фостер (2007) сделал вывод, что перья Ornitholestes, вероятно, были более примитивными, чем перья птиц; они, вероятно, «покрыли бы все тело, за исключением ног, коротким пальто, возможно, с более длинными перьями, выстилающими верхнюю часть черепа или шеи и задний край предплечий». Если предположить, что они присутствовали, эти перья могли использоваться для изоляции и, возможно, также для высиживания яиц.
| year =()| year =()| year =()| year =()
