
Войти
 Берлинский образец Archeopteryx lithographica
Берлинский образец Archeopteryx lithographica Научный вопрос, в пределах какой группы животных птиц Evolved традиционно называли «происхождением птиц ». В настоящее время научный консенсус состоит в том, что птицы представляют собой группу манирапторанов теропод динозавров, которые произошли в мезозойскую эру.
Тесная связь между птицами и динозаврами была впервые предложена в девятнадцатом веке после открытия примитивной птицы археоптерикс в Германии. Птицы и вымершие динозавры, не являющиеся птицами, имеют много общих черт скелета. Кроме того, были собраны окаменелости более тридцати видов нептичьих динозавров с сохранившимися перьями. Есть даже очень маленькие динозавры, такие как Microraptor и Anchiornis, у которых длинные, лопастные перья на руках и ногах образуют крылья. В юрском базальном слое avialan Pedopenna также показаны эти длинные перья на ступнях. Палеонтолог Лоуренс Уитмер в 2009 году пришел к выводу, что этих свидетельств достаточно, чтобы продемонстрировать, что эволюция птиц прошла стадию с четырьмя крыльями. Ископаемые останки также демонстрируют, что птицы и динозавры имеют общие черты, такие как полые, пневматизированные кости, гастролиты в пищеварительной системе, строение гнезд и наседка..
Хотя происхождение птиц исторически спорный вопрос в эволюционная биология, лишь немногие ученых до сих пор спорят о динозавровом происхождении птиц, предлагая происхождение от других видов archosaurian рептилии. В рамках консенсуса, который поддерживает происхождение динозавров, точная последовательность эволюционных событий, которые привели к появлению ранних птиц в пределах теропод-манирапторан, оспаривается. Происхождение птичьего полета - отдельный, но связанный вопрос, на который также есть несколько предложенных ответов.
 Томас Генри Хаксли (1825–1895)
Томас Генри Хаксли (1825–1895) Научный расследование происхождения птиц началось вскоре после публикации в 1859 году книги Чарльза Дарвина О происхождении видов. В 1860 г. окаменелое перо было обнаружено в Германии поздней юре известняке Solnhofen. Христиан Эрих Герман фон Мейер в следующем году описал это перо как Archeopteryx lithographica. Ричард Оуэн описал почти полный скелет в 1863 году, признав его птицей, несмотря на многие черты, напоминающие рептилии, включая когтистые передние конечности и длинный костлявый хвост.
Биолог Томас Генри Хаксли, известный как «Бульдог Дарвина» за его упорную поддержку новой теории эволюции средствами естественного отбора, почти сразу схватили археоптерикс как переходное ископаемое между птицами и рептилиями. Начиная с 1868 года и следуя более ранним предложениям Карла Гегенбаура и Эдварда Дринкера Коупа, Хаксли провел подробное сравнение археоптерикса с различными доисторическими рептилиями и обнаружил, что он больше всего похож на динозавров, таких как Гипсилофодон и Компсогнат. Открытие в конце 1870-х годов культового «берлинского экземпляра» археоптерикса с целым набором зубов рептилии дало дополнительные доказательства. Как и Коп, Хаксли предложил эволюционную связь между птицами и динозаврами. Хотя Хаксли был противником очень влиятельного Оуэна, его выводы были приняты многими биологами, включая барона Франца Нопса, в то время как другие, в частности Гарри Сили, утверждали, что сходство было связано с конвергентная эволюция.
Поворотный момент наступил в начале двадцатого века с работами Герхарда Хейльмана из Дании. Художник по профессии, Хейльманн проявлял научный интерес к птицам и с 1913 по 1916 год, развивая более ранние работы Отенио Абеля, опубликовал результаты своих исследований в нескольких частях, касающихся анатомии, эмбриология, поведение, палеонтология и эволюция птиц. Его работа, первоначально написанная на датском как Vor Nuvaerende Viden om Fuglenes Afstamning, была собрана, переведена на английский и опубликована в 1926 году как Происхождение птиц.
 Гипотетическая иллюстрация Хейльмана пары боевой 'Proaves ' от 1916 года
Гипотетическая иллюстрация Хейльмана пары боевой 'Proaves ' от 1916 года Подобно Хаксли, Хейльманн сравнил археоптерикс и других птиц с исчерпывающим списком доисторических рептилий, а также пришел к выводу, что динозавры-теропод, такие как Компсогнат, были наиболее похожи. Однако Хейльманн отметил, что у птиц были ключицы (ключицы), сросшиеся с образованием кости, называемой furcula («поперечный рычаг»), и хотя ключицы были известны у более примитивных рептилий, они имели еще не обнаружен у динозавров. Поскольку он твердо верил в закон Долло, который гласит, что эволюция необратима, Хейльманн не мог согласиться с тем, что ключицы были потеряны у динозавров и повторно эволюционировали у птиц. Поэтому он был вынужден исключить динозавров как предков птиц и приписать все их сходства конвергентной эволюции. Хейльманн заявил, что предки птиц вместо этого будут найдены среди более примитивных рептилий "текодонта ". Благодаря чрезвычайно тщательному подходу Хейльмана его книга стала классикой в этой области, а его выводы о происхождении птиц, как и по большинству других тем, были приняты почти всеми эволюционными биологами в течение следующих четырех десятилетий.
Ключицы относительно относительно. хрупкие кости и, следовательно, могут быть разрушены или, по крайней мере, повреждены до неузнаваемости. Тем не менее, некоторые ископаемые ключицы теропод были раскопаны до того, как Хейльман написал свою книгу, но они были неправильно идентифицированы. Отсутствие ключиц у динозавров стало ортодоксальной точкой зрения, несмотря на открытие ключиц у примитивных теропод Сегизавр в 1936 году. Следующее сообщение о ключицах у динозавров было в российской статье 1983 года.
Вопреки тому, во что верил Хейльманн, палеонтологи теперь признают, что ключицы и в большинстве случаев фуркулы являются стандартной чертой не только теропод, но и заурисхических динозавров. Вплоть до конца 2007 г. окостеневшие фуркулы (т.е. сделанные из кости, а не хряща ) были обнаружены у всех типов теропод, кроме самых базальных, Eoraptor и Herrerasaurus. Первоначальное сообщение о фуркуле у примитивного теропод Segisaurus (1936) было подтверждено повторным исследованием в 2005 году. Сочлененные, похожие на фуркулу ключицы были также обнаружены у Massospondylus, Ранняя юра зауроподоморф.
 Сходство передних конечностей Дейнониха (слева) и археоптерикса (справа) привело Джон Остром, чтобы восстановить связь между динозаврами и птицами.
Сходство передних конечностей Дейнониха (слева) и археоптерикса (справа) привело Джон Остром, чтобы восстановить связь между динозаврами и птицами. Тенденция начала разворачиваться против гипотезы «текодонтов» после открытия в 1964 году нового динозавра-теропод в Монтане. В 1969 году этот динозавр был описан и назван Дейнонихом Джоном Остромом из Йельского университета. В следующем году Остром повторно описал экземпляр Pterodactylus в голландском музее Тейлера как еще один скелет археоптерикса. Образец состоял в основном из одного крыла, и его описание заставило Острома понять сходство между запястьями Археоптерикса и Дейнониха.
В 1972 году британский палеонтолог Алик Уокер предположил, что птицы произошли не от «текодонтов», а от крокодилов предков, таких как Sphenosuchus. Работа Острома как с тероподами, так и с ранними птицами побудила его ответить серией публикаций в середине 1970-х годов, в которых он изложил многие сходства между птицами и тероподными динозаврами, возродив идеи, впервые высказанные Хаксли более века назад. Признание Остромом динозавров предков птиц, наряду с другими новыми идеями о метаболизме динозавров, уровнях активности и родительской заботе, положило начало тому, что известно как ренессанс динозавров, который начался в 1970-х годах и продолжается по сей день..
Откровения Острома также совпали с растущим принятием филогенетической систематики (кладистика ), которое началось в 1960-х годах с работы Вилли Хеннига. Кладистика - это точный метод систематизации видов, основанный строго на их эволюционных отношениях, которые рассчитываются путем определения эволюционного древа, подразумевающего наименьшее количество изменений в их анатомических характеристиках. В 1980-х годах кладистическая методология была впервые применена к филогении динозавров Жаком Готье и другими, недвусмысленно показав, что птицы были производной группой динозавров-теропод. Ранние исследования показали, что тероподы дромеозаврид, такие как Deinonychus, были особенно тесно связаны с птицами, и этот результат с тех пор неоднократно подтверждался.
 Ископаемые остатки Sinosauropteryx prima
Ископаемые остатки Sinosauropteryx prima В начале 1990-х гг. обнаружение прекрасно сохранившихся окаменелостей птиц в нескольких раннемеловых геологических формациях в северо-восточной китайской провинции Ляонин. В 1996 году китайские палеонтологи описали Sinosauropteryx как новый род птиц из формации Yixian, но это животное было быстро признано более базальным динозавром-теропод, близким к Compsognathus. Удивительно, но его тело было покрыто длинными нитевидными структурами. Их окрестили «праоперями» и сочли гомологичными более развитым перьям птиц, хотя некоторые ученые не согласны с этой оценкой. Китайские и североамериканские ученые вскоре описали Caudipteryx и Protarchaeopteryx. Судя по особенностям скелета, эти животные не были птичьими динозаврами, но их останки имели полностью сформированные перья, очень похожие на перья птиц. «Археораптор », описанный без экспертной оценки в выпуске журнала National Geographic за 1999 год, оказался контрабандным подделкой, но законные останки продолжают выливаться из иссянь, как легально, так и незаконно. Перья, или «праоперья», были обнаружены у самых разных теропод в исяне, и открытие чрезвычайно похожих на птиц нептичьих динозавров, а также нептичьих динозавроподобных примитивных птиц почти полностью закрыло морфологический пробел. между нептичьими тероподами и птицами.
Между эмбриологами и палеонтологами ведутся споры о том, действительно ли руки теропод динозавров и птиц разные, основанные на подсчете фаланг, подсчете количества фаланг (пальцев) в руке. Это важная и горячо обсуждаемая область исследования, поскольку ее результаты могут поставить под сомнение консенсус в отношении того, что птицы являются (потомками) динозавров.
Эмбриологи и некоторые палеонтологи, выступающие против связи между птицами и динозаврами, давно пронумеровали пальцы птиц II-III-IV на основании многочисленных исследований развития в яйце. Это основано на том факте, что в большинстве амниот первая цифра, которая образуется в пятипалой руке, - это цифра IV, которая образует первичную ось. Таким образом, эмбриологи определили первичную ось у птиц как палец IV, а выжившие пальцы - как II-III-IV. Окаменелости передовых рук теропод (Tetanurae ), по-видимому, имеют пальцы I-II-III (некоторые роды в пределах Avetheropoda также имеют уменьшенный палец IV). Если это правда, то развитие пальцев у птиц на II-III-IV является показателем против происхождения теропод (динозавров). Однако при отсутствии онтогенных (онтогенетических) оснований для окончательного определения, какие пальцы какие на руке теропод (поскольку сегодня невозможно наблюдать рост и развитие нептичьих теропод), маркировка руки теропод не является абсолютно убедительно.
Палеонтологи традиционно идентифицировали пальцы птиц как I-II-III. Они утверждают, что пальцы птиц имеют номера I-II-III, как и у тероподных динозавров, согласно законсервированной формуле фаланг. Количество фалангов у архозавров - 2-3-4-5-3; многие родословные архозавров имеют уменьшенное количество цифр, но имеют ту же формулу фаланги в оставшихся цифрах. Другими словами, палеонтологи утверждают, что архозавры разных родов имеют тенденцию терять одни и те же пальцы, когда происходит потеря пальца, снаружи внутрь. Три пальца дромеозавров и археоптерикс имеют ту же фаланговую формулу I-II-III, что и пальцы I-II-III базальных архозавров. Следовательно, потеряны цифры V и IV. Если это правда, то современные птицы также будут обладать I-II-III пальцами. Кроме того, в одной публикации 1999 г. было предложено сдвинуть рамки цифр линии теропод, ведущих к птицам (таким образом, превратив цифру I в цифру II, цифру II в цифру III и т. Д.). Однако такие сдвиги скелета редки у амниот и - чтобы соответствовать тероподному происхождению птиц - должны были произойти исключительно в передних конечностях линии птица-теропод, а не в задних конечностях (состояние, неизвестное ни у одного животного). Это называется латеральным сокращением цифр (LDR) по сравнению с двусторонним сокращением цифр (BDR) (см. Также Limusaurus
Небольшое меньшинство, известное под аббревиатурой BAND (птицы - не динозавры), включая орнитологов Алан Федучча и Ларри Мартин продолжают утверждать, что птицы более тесно связаны с более ранними рептилиями, такими как Longisquama или Euparkeria, чем с Эмбриологические исследования птиц биология развития подняли вопросы о гомологии пальцев в передних конечностях птиц и динозавров. Однако из-за убедительных доказательств, предоставленных сравнительной анатомией и филогенетикой, а также драматические окаменелости пернатых динозавров из Китая, идея о том, что птицы являются производными динозаврами, сначала отстаиваемая Хаксли, а затем Нопса и Остром, пользуется почти единодушной поддержкой среди современных палеонтологов.
В публикации 2011 г. было предложено выбрать расширение скелета Движущей силой появления этого клада была не эволюция полета, а мускул. Согласно этой гипотезе, мышцы у предполагаемых эндотермических ящеров стали больше в ответ на потерю митохондрий позвоночных разобщающий белок, UCP1, который является термогенным. У млекопитающих UCP1 функционирует в коричневой жировой ткани для защиты новорожденных от гипотермии. У современных птиц скелетные мышцы выполняют аналогичную функцию и, как предполагается, выполняли это у их предков. С этой точки зрения, двуногие и другие изменения скелета птиц были побочными эффектами мышечной гиперплазии с дальнейшими эволюционными модификациями передних конечностей, включая адаптацию к полету или плаванию, и рудиментарность, являющаяся вторичным следствием двуногости.
Исторически археоптерикс считался первой птицей, или урвогелем. Хотя более новые открытия окаменелостей заполнили пробел между тероподами и археоптериксом, а также пропасть между археоптериксом и современными птицами, филогенетические систематики, следуя традиции, почти всегда используют археоптерикса в качестве спецификатора, чтобы помочь определить Aves. Реже авес определялся как кроновая группа, состоящая только из современных птиц. Почти все палеонтологи считают птиц целурозаврами теропод динозаврами. Внутри целурозаврии многочисленные кладистические анализы подтвердили наличие клады под названием Maniraptora, состоящей из теризинозавроидов, овирапторозавров, троодонтиды, дромеозавриды и птицы. Из них дромеозавриды и троодонтиды обычно объединяются в кладу Deinonychosauria, которая является сестринской группой для птиц (вместе формируя кладу-узел Eumaniraptora ) внутри stem-clade Paraves.
В других исследованиях предложены альтернативные филогении, в которых определенные группы динозавров, обычно считающиеся нептичьими, могли развиться от птичьих предков. Например, анализ 2002 года показал, что овирапторозавры были базальными птицами. Альваресзавриды, известные из Азии и Америки, по-разному классифицируются как базальные манирапторы, паравианцы, сестринский таксон орнитомимозавров, а также специализированные ранние птицы. Род Rahonavis, первоначально описанный как ранняя пташка, в нескольких исследованиях был идентифицирован как нептичий дромеозаврид. Было высказано предположение, что сами дромеозавриды и троодонтиды находятся внутри Авеса, а не вне его.
Многие анатомические особенности характерны для птиц и теропод. динозавры.
Археоптерикс, первый хороший пример «пернатого динозавра», был обнаружен в 1861 году. Первый образец был обнаружен в известняке в Зольнхофене на юге Германии, который представляет собой lagerstätte, редкое и замечательное геологическое образование, известное своими великолепно детализированными окаменелостями. Археоптерикс - это переходное ископаемое, с чертами, явно промежуточными между признаками нептичьих тероподовых динозавров и птиц. Обнаруженный всего через два года после основополагающей работы Дарвина Происхождение видов, его открытие спровоцировало зарождающиеся дебаты между сторонниками эволюционной биологии и креационизма. Эта ранняя пташка настолько похожа на динозавра, что без четкого отпечатка перьев в окружающей скале, по крайней мере, один экземпляр был ошибочно принят за Compsognathus.
 Части пера
Части пера С 1990-х годов появилось несколько дополнительных пернатых динозавров, что является еще более убедительным доказательством тесной связи между динозаврами и современными птицами. Первые из них первоначально были описаны как простые нитевидные протоиери, о которых сообщалось в родословных динозавров столь же примитивных, как компсогнатиды и тираннозавроиды. Однако перья, неотличимые от перьев современных птиц, вскоре были обнаружены и у нептичьих динозавров.
Незначительное меньшинство исследователей заявили, что простые нитевидные структуры «прото-перья» являются просто результатом разложения перьев. коллагеновые волокна под кожей динозавров или в плавниках вдоль их спины, и те виды с бесспорными перьями, как овирапторозавры и дромеозавры, не динозавры, а настоящие птицы, не связанные с динозаврами. Однако большинство исследований пришли к выводу, что пернатые динозавры на самом деле являются динозаврами, и что более простые волокна бесспорных теропод представляют собой простые перья. Некоторые исследователи продемонстрировали наличие цветосодержащего меланина в структурах, чего можно было бы ожидать в перьях, но не в коллагеновых волокнах. Другие продемонстрировали, используя исследования разложения современных птиц, что даже развитые перья кажутся нитевидными, когда подвергаются сокрушающим силам, испытываемым во время окаменелости, и что предполагаемые «праоперья», возможно, были более сложными, чем считалось ранее. Детальное исследование «праперья» Sinosauropteryx prima показало, что отдельные перья состояли из центрального пера (рахиса) с отходящими от него более тонкими зазубринами, похожими на перья современных птиц, но более примитивными по структуре.
 Ископаемые остатки NGMC 91, вероятный образец синорнитозавра
Ископаемые остатки NGMC 91, вероятный образец синорнитозавра Поскольку перья часто ассоциируются с птицами, пернатые динозавры часто рекламируются как недостающее звено между птицами и динозаврами. Однако многочисленные особенности скелета, общие для обеих групп, представляют собой более важную связь для палеонтологов. Кроме того, становится все более очевидным, что отношения между птицами и динозаврами и эволюция полета - более сложные темы, чем предполагалось ранее. Например, хотя когда-то считалось, что птицы эволюционировали от динозавров в одну линейную последовательность, некоторые ученые, в первую очередь Грегори С. Пол, пришли к выводу, что динозавры, такие как дромеозавры, могли эволюционировать от птиц, теряющих способность летать, сохраняя при этом свои перья, как у современных страусиных и других ратитов.
Сравнение скелетов птиц и динозавров, а также кладистический анализ, усиливает аргументы в пользу связи, особенно в отношении ветви теропод, называемой манирапторами. Скелетные сходства включают шею, лобок, запястье (полулунное запястное ), руку и грудной пояс, лопатку., ключица и грудная кость.
Исследование, сравнивающее черепа эмбрионов, молодых и взрослых архозавров, пришло к выводу, что черепа птиц произошли от черепа теропод динозавров прогенез, тип педоморфной гетерохронии, в результате которой сохраняются ювенильные характеристики их предков.
 Сравнение воздушных мешков маджунгазавра и птицы (утка)
Сравнение воздушных мешков маджунгазавра и птицы (утка) Согласно исследованию, проведенному Патриком М. О'Коннором из Университета Огайо, у крупных динозавров, питающихся мясом, была сложная система воздушных мешочков, подобных тем, которые встречаются у современных птиц. У динозавров-теропод (плотоядные животные, которые ходили на двух ногах и имели птицеподобные лапы) гибкие воздушные мешочки из мягких тканей, вероятно, перекачивали воздух через жесткие легкие, как в случае с птицами. «То, что когда-то формально считалось уникальным для птиц, в той или иной форме присутствовало у предков птиц», - сказал О'Коннор.
Компьютерная томография (КТ), проведенная в 2000 г. в грудной полости образца орнитопода тесцелозавра были обнаружены очевидные остатки сложного четырехкамерного сердца, очень похожего на те, что обнаруживаются у современных млекопитающих и птиц. Идея вызывает споры в научном сообществе, критикуется за плохую анатомию или просто принятие желаемого за действительное.
В исследовании, опубликованном в 2011 году, к вопросу о личности объекта использовалось несколько направлений исследования, включая более продвинутое компьютерное сканирование, гистология, дифракция рентгеновских лучей, рентгеновская фотоэлектронная спектроскопия и сканирующая электронная микроскопия. С помощью этих методов авторы обнаружили, что: внутренняя структура объекта не включает камеры, а состоит из трех несвязанных областей материала с более низкой плотностью и не сопоставима со структурой сердца страуса ; «стены» состоят из осадочных минералов, которые, как известно, не образуются в биологических системах, таких как гетит, полевой шпат минералы, кварц и гипс, а также некоторые фрагменты растений; углерод, азот и фосфор, химические элементы, важные для жизни, отсутствовали в их образцах; клеточные структуры сердца отсутствовали. Был один возможный участок с клеточными структурами животных. Авторы обнаружили, что их данные подтверждают идентификацию как конкрецию песка из погребальной среды, а не сердца, с возможностью сохранения отдельных участков тканей.
Вопрос о том, как эта находка отражает скорость метаболизма и внутреннее состояние динозавра. анатомия, однако, спорный вопрос, независимо от идентичности объекта. У современных крокодилов и птиц, ближайших ныне живущих родственников динозавров, есть четырехкамерные сердца (хотя и модифицированные у крокодилов), так что динозавры, вероятно, тоже имели их; структура не обязательно связана со скоростью метаболизма.
Окаменелости троодонтов Mei и Sinornithoides демонстрируют что динозавры спали, как некоторые современные птицы, закрыв головы под мышками. Такое поведение, которое могло помочь согреть голову, также характерно для современных птиц.
При откладывании яиц у самок птиц вырастают особые кости в конечностях. Эта медуллярная кость образует богатый кальцием слой внутри твердой внешней кости и используется в качестве источника кальция для изготовления яичной скорлупы. Присутствие эндостальных костных тканей, выстилающих внутренние полости костного мозга частей задней конечности экземпляра Tyrannosaurus rex, позволяет предположить, что T. rex использовал аналогичные репродуктивные стратегии, и показало, что образец - самка. Дальнейшие исследования обнаружили костный мозг теропод Allosaurus и орнитопод Tenontosaurus. Поскольку линия динозавров, которая включает аллозавров и тираннозавров, расходилась с линией, которая привела к тенонтозавру на очень раннем этапе эволюции динозавров, это предполагает, что динозавры в целом производили костную ткань.
 Гнездящийся экземпляр Citipati osmolskae в Американском музее естественной истории в Нью-Йорке
Гнездящийся экземпляр Citipati osmolskae в Американском музее естественной истории в Нью-Йорке Несколько экземпляров Citipati были найдены покоящимися над яйца в гнезде в положении, наиболее напоминающем выводок.
. Многочисленные виды динозавров, например Maiasaura, были обнаружены в стадах, смешанных как с очень молодыми, так и с взрослыми особями, что свидетельствует о тесном взаимодействии между ними.
Эмбрион динозавра был найден без зубов, что указывает на необходимость некоторой родительской заботы, чтобы накормить молодого динозавра, возможно, взрослый динозавр срыгнул пищу в рот молодого динозавра (см. altricial ). Такое поведение наблюдается у многих видов птиц; родительские птицы срыгивают пищу в рот птенца.
И птицы, и динозавры используют мускульные камни. Эти камни глотаются животными, чтобы помочь пищеварению и расщеплению пищи и твердых волокон, когда они попадают в желудок. При обнаружении вместе с окаменелостями камни в желудке называются гастролитами. Камни желудочно-кишечного тракта также встречаются у некоторых рыб (кефаль, грязь и гиллару, разновидность форели) и крокодилов.
В нескольких случаях заявлялось об извлечении ДНК и белков из окаменелостей мезозойских динозавров, что позволяет проводить сравнение с птицами. Несколько белков предположительно были обнаружены в окаменелостях динозавров, в том числе гемоглобин.
В мартовском выпуске журнала Science за 2005 год, доктор Мэри Хигби Швейцер и ее команда объявила об открытии гибкого материала, напоминающего настоящие мягкие ткани, внутри Tyrannosaurus rex ноги кости образца MOR 1125 из формации Хелл-Крик в Монтана. Семь типов коллагена, полученных из костных фрагментов, по сравнению с данными о коллагене у живых птиц (в частности, цыпленка ), позволяют предположить, что более старые тероподы и птицы тесно связаны. Мягкие ткани позволили провести молекулярное сравнение клеточной анатомии и секвенирование белка ткани коллаген, опубликованное в 2007 году, оба из которых показали, что T. rex и птицы больше тесно связаны друг с другом, чем с Аллигатором. Второе молекулярное исследование убедительно подтвердило связь птиц с динозаврами, хотя оно не поместило птиц в теропод, как ожидалось. В этом исследовании использовали восемь дополнительных последовательностей коллагена, извлеченных из бедренной кости "мумифицированного" Brachylophosaurus canadensis образца MOR 2598, гадрозавра. Однако эти результаты были очень противоречивыми. О других пептидах мезозойского возраста не сообщалось. В 2008 году было высказано предположение, что предполагаемая мягкая ткань на самом деле является бактериальной микропленкой. В ответ было заявлено, что именно эти микропленки защищают мягкие ткани. Другое возражение заключалось в том, что результаты могли быть вызваны заражением. В 2015 году в более контролируемых условиях, обеспечивающих защиту от заражения, пептиды все же были идентифицированы. В 2017 году исследование показало, что в костях современного страуса присутствует пептид, идентичный таковому у образцов тираннозавра и брахилофозавра, что подчеркивает опасность перекрестного заражения.
Успешная экстракция О древней ДНК из окаменелостей динозавров сообщалось в двух разных случаях, но после дальнейшего изучения и экспертной оценки ни одно из этих сообщений не могло быть подтверждено.
Споры о происхождении птичьего полета почти так же стары, как идея о том, что птицы произошли от динозавров, которая возникла вскоре после открытия археоптерикса в 1862 году. В большинстве случаев доминировали две теории. обсуждение с тех пор: беглая («с нуля») теория предполагает, что птицы произошли от маленьких, быстрых хищников, которые бегали по земле; Древесная («с деревьев вниз») теория предполагает, что полет с приводом от двигателя произошел от планирования без двигателя древесных (лазающих по деревьям) животных. Более поздняя теория, «бег по наклонной поверхности с использованием крыльев» (WAIR), является вариантом бегущей теории и предполагает, что крылья развили свои аэродинамические функции в результате необходимости быстро бегать по очень крутым склонам. такие как деревья, которые помогли бы маленьким пернатым динозаврам убежать от хищников.
В марте 2018 года ученые сообщили, что археоптерикс, вероятно, был способен летать, но в манере, существенно отличавшейся от современных птиц.
 Реконструкция Рахонависа, наземного пернатого динозавра, который, по мнению некоторых исследователей, был хорошо оборудован для полета
Реконструкция Рахонависа, наземного пернатого динозавра, который, по мнению некоторых исследователей, был хорошо оборудован для полета Беглая теория происхождения полета была первой предложено Сэмюэлем Венделлом Уиллистоном и развито бароном Нопса. Эта гипотеза предполагает, что некоторые быстро бегающие животные с длинными хвостами использовали свои руки, чтобы сохранять равновесие во время бега. Современные версии этой теории во многих деталях отличаются от версии Уиллистона-Нопса, главным образом в результате открытий со времен Нопсы.
Нопса предположил, что увеличение площади вытянутых рук могло помочь маленьким бегающим хищникам сохранять равновесие, а чешуя предплечий удлинялась, превращаясь в перья. Перья также могли использоваться для ловли насекомых или другой добычи. Постепенно животные прыгали на большие расстояния, чему способствовали их развивающиеся крылья. Нопса также предложил три стадии эволюции полета. Во-первых, у животных развился пассивный полет, при котором развивающиеся структуры крыльев служили своего рода парашютом. Во-вторых, они достигли активного полета за счет взмахов крыльев. Он использовал археоптерикс в качестве примера этой второй стадии. Наконец, птицы обрели способность парить.
 Предлагаемое развитие полета в книге 1922 года: Тетраптерикс, Археоптерикс, Гипотетическая стадия, Современная птица
Предлагаемое развитие полета в книге 1922 года: Тетраптерикс, Археоптерикс, Гипотетическая стадия, Современная птица Текущее мнение таково, что перья не произошли из чешуи, поскольку перья состоят из разные белки. Более серьезно, теория Нопса предполагает, что перья эволюционировали как часть эволюции полета, и недавние открытия доказывают, что это предположение неверно.
Перья очень распространены у целурозавров (включая раннего тираннозавроида Дилонг ). Современные птицы классифицируются как целурозавры почти всеми палеонтологами, но не некоторыми орнитологами. Современная версия гипотезы «с нуля» утверждает, что предки птиц были небольшими пернатыми хищными динозаврами, бегающими по земле (скорее похожими на дорожных бегунов в их стиле охоты), которые использовали свои передние конечности для равновесия при преследовании жертва, и что передние конечности и перья позже эволюционировали таким образом, что они обеспечивали планирование, а затем и движение. Наиболее широко предполагаемые первоначальные функции перьев включают теплоизоляцию и конкурентоспособность, как у современных птиц.
Все окаменелости археоптериксов происходят из морских отложений, и было высказано предположение, что крылья могли помочь птицам перебегать вода в манере Ящерицы Иисуса Христа (василиск обыкновенный ).
Самые последние опровержения гипотезы «с нуля» - попытка опровергнуть современную версию предположения, что птицы являются модифицированными целурозаврами динозавров. Самые сильные атаки основаны на эмбриологический анализ, в котором делается вывод, что крылья птиц образуются из цифр 2, 3 и 4 (что соответствует указательному, среднему и безымянному пальцам человека. Первый из трех пальцев птицы образует alula, которую они используют, чтобы избежать сваливания в полете на малой скорости - например, при приземлении). Однако руки целурозавров образованы цифрами 1, 2 и 3 (большой палец и первые два пальца у человека). Однако эти эмбриологические анализы ses были немедленно оспорены на том эмбриологическом основании, что «рука» часто развивается по-другому в кладах, которые потеряли несколько пальцев в ходе своей эволюции, и что «руки» птиц действительно развиваются из пальцев 1, 2, и 3. Эта дискуссия сложна и еще не решена - см. «Гомология цифр ».
Гипотеза бег по наклонной поверхности с крыльями (WAIR) была вызвана наблюдениями за молодыми цыплятами чукар и предлагает эти крылья приобрели свои аэродинамические функции в результате необходимости быстро подниматься по очень крутым склонам, таким как стволы деревьев, например, чтобы убежать от хищников. Это делает ее специализированным типом беглой («с нуля») теории. Обратите внимание, что в этом сценарии птицам требуется прижимная сила, чтобы ноги лучше держались за них. Но у ранних птиц, включая археоптерикса, отсутствовал плечевой механизм, с помощью которого крылья современных птиц производили быстрые и мощные взмахи вверх. Поскольку прижимная сила WAIR, от которой зависит, создается восходящими ходами, кажется, что ранние пташки были неспособны к WAIR. Поскольку WAIR - это поведенческая черта без остеологической специализации, филогенетическое размещение взлетающего гребня перед расхождением Neornithes, группы, которая включает всех существующих птиц, делает невозможным определение того, является ли WAIR предком птичьего полетный удар или производный от него.
 Четырехкрылый Микрораптор, член группы Dromaeosauridae, близкородственных динозавров птицам
Четырехкрылый Микрораптор, член группы Dromaeosauridae, близкородственных динозавров птицам Большинство версий древесной гипотезы утверждают, что предки птиц были очень маленькими динозаврами, которые жили на деревьях, прыгая с ветки на ветку. У этого маленького динозавра уже были перья, которые эволюция использовала для создания более длинных и жестких форм, которые использовались в аэродинамике, в конечном итоге создавая крылья. Затем крылья развивались и становились все более совершенными в качестве устройств, позволяющих прыгунам больше контролировать, прыгать с парашютом, парить и ступенчато летать. Древесная гипотеза также отмечает, что для древесных животных аэродинамика гораздо более энергоэффективна, поскольку такие животные просто падают, чтобы достичь минимальной скорости планирования.
Несколько маленьких динозавров юрского или раннего мелового периода, все с перьями, имели были интерпретированы как возможно имеющие древесные и / или аэродинамические приспособления. К ним относятся Scansoriopteryx, Epidexipteryx, Microraptor, Pedopenna и Anchiornis. Anchiornis особенно важен для этого предмета, так как он жил в начале поздней юры, задолго до археоптерикса.
Анализ пропорций костей пальцев самых примитивных птиц, археоптерикса и Confuciusornis по сравнению с таковыми у живых видов, предполагают, что ранние виды могли жить как на земле, так и на деревьях.
Одно исследование показало, что самые ранние птицы и их непосредственные предки не лазили по деревьям. Это исследование показало, что степень искривления когтей на лапах ранних птиц была больше похожа на ту, что наблюдается у современных наземных птиц, чем у птиц-наседов.
археоптерикс был первым и долгое время единственное известное пернатое мезозойское животное. В результате обсуждение эволюции птиц и их полета сосредоточилось на археоптериксе, по крайней мере, до середины 1990-х годов.
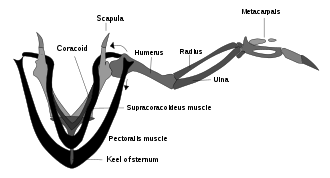 Supracoracoideus работает, используя подобную шкиву систему, чтобы поднять крыло, в то время как грудные мышцы обеспечивают мощный толчок вниз.
Supracoracoideus работает, используя подобную шкиву систему, чтобы поднять крыло, в то время как грудные мышцы обеспечивают мощный толчок вниз. Были споры о том, действительно ли археоптерикс может летать. Похоже, что у археоптерикса были мозговые структуры и датчики баланса внутреннего уха, которые птицы используют для управления своим полетом. У археоптерикса также было расположение перьев крыльев, как у современных птиц, и такие же асимметричные маховые перья на крыльях и хвосте. Но у археоптерикса отсутствовал плечевой механизм, с помощью которого крылья современных птиц производили быстрые и мощные взмахи вверх (см. Диаграмму выше супракоракоидного шкива); это может означать, что он и другие ранние птицы были неспособны к взмаху крыльев и могли только парить.
Но открытие с начала 1990-х годов многих пернатых динозавров означает, что археоптерикс больше не является ключевым фигура в эволюции птичьего полета. Другие небольшие пернатые целурозавры из мела и поздней юры показывают возможные предшественники птичьего полета. К ним относятся Рахонавис, наземный бегун с поднятым серпоподобным когтем на втором пальце ноги, похожим на велоцираптора, который, по мнению некоторых палеонтологов, был лучше приспособлен для полета, чем археоптерикс, Скансориоптерикс, древесный динозавр, который может поддерживать теорию «с деревьев вниз», и Микрораптор, древесный динозавр, возможно, способный летать на приводе, но если так, то больше похож на биплан, так как у него были хорошо развитые перья на ногах. Еще в 1915 году некоторые ученые утверждали, что эволюция полета птиц могла пройти стадию четырехкрылости (или тетраптерикса).
| Целурозавры |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Гипотеза, приписываемая Грегори Полу и выдвинутая в его книгах «Хищные динозавры мира» (1988) и «Динозавры» of the Air (2002), предполагает, что некоторые группы нелетающих хищных динозавров - особенно дейнонихозавры, но, возможно, другие, такие как овирапторозавры, теризинозавры, альвареззавриды и орнитомимозавры - фактически произошли от птиц. Пол также предположил, что птица-предок этих групп была более развита в своих летных адаптациях, чем археоптерикс. Гипотеза означала бы, что археоптерикс менее связан с существующими птицами, чем эти динозавры.
Гипотеза Пола получила дополнительную поддержку, когда Mayr et al. (2005) проанализировали новый, десятый образец археоптерикса и пришли к выводу, что археоптерикс был сестринской кладой по отношению к дейнонихозаврам, но что более продвинутая птица Confuciusornis принадлежала к Dromaeosauridae. Этот результат подтверждает гипотезу Пола, предполагающую, что Deinonychosauria и Troodontidae являются частью Aves, собственно птичьего происхождения и, во-вторых, нелетающими. В этот документ, однако, были исключены все другие птицы, и поэтому не проводилась выборка распределения их признаков. Статья подверглась критике со стороны Корфа и Батлера (2006), которые обнаружили, что авторы не могут подтвердить свои выводы статистически. Mayr et al. согласились с тем, что статистическая поддержка была слабой, но добавила, что она также слаба для альтернативных сценариев.
Текущий кладистический анализ не поддерживает гипотезу Пола о положении археоптерикса. Вместо этого они указывают на то, что археоптерикс ближе к птицам в кладе Avialae, чем к дейнонихозаврам или овирапторозаврам. Однако некоторые окаменелости подтверждают версию этой теории, согласно которой у некоторых нелетающих хищных динозавров могли быть летающие предки. В частности, Microraptor, Pedopenna и Anchiornis имеют крылатые лапы, имеют много общих черт и лежат близко к основанию клады Paraves. Это говорит о том, что предком паравианца был четырехкрылый планер, и что более крупные дейнонихозавры вторично утратили способность к полету, в то время как аэродинамические способности птичьего происхождения увеличивались по мере развития. Дейноних может также демонстрировать частичную подвижность, с молодые существа способны летать или планировать, а взрослые - нелетающие. В 2018 году исследование пришло к выводу, что у последнего общего предка Pennaraptora были суставные поверхности на пальцах и между плюсневой костью и запястьем, которые были оптимизированы для стабилизации руки в полете. Это рассматривалось как признак вторичной нелетаемости у тяжелых базальных представителей этой группы.
В Euornithes самым ранним однозначным примером вторичной нелетаемости является Patagopteryx.
![]() Портал динозавров
Портал динозавров
| На Wikimedia Commons есть материалы, связанные с . окаменелостями Авеса. |