
Войти
 Рабочие пчелы добывают нектар не только для себя, но и для всего своего ульевого сообщества. Теория оптимального кормодобывания предсказывает, что эта пчела будет кормиться таким образом, чтобы максимизировать чистый выход энергии из своего улья.
Рабочие пчелы добывают нектар не только для себя, но и для всего своего ульевого сообщества. Теория оптимального кормодобывания предсказывает, что эта пчела будет кормиться таким образом, чтобы максимизировать чистый выход энергии из своего улья. Теория оптимального кормодобывания (OFT ) - это поведенческая экология модель, которая помогает предсказать, как животное ведет себя при поиске пищи. Хотя добыча пищи дает животному энергию, поиск и захват пищи требуют как энергии, так и времени. Чтобы максимизировать приспособленность, животное использует стратегию кормодобывания, которая обеспечивает наибольшую выгоду (энергию) при наименьших затратах, максимизируя полученную чистую энергию. OFT помогает предсказать наилучшую стратегию, которую животное может использовать для достижения этой цели.
OFT - это экологическое приложение модели оптимальности . Эта теория предполагает, что наиболее экономически выгодный режим кормодобывания будет выбран для вида посредством естественного отбора. Говорят, что при использовании OFT для моделирования пищевого поведения организмы максимизируют переменную, известную как валюта, например, наибольшее количество пищи в единицу времени. Кроме того, ограничения среды - это другие переменные, которые необходимо учитывать. Ограничения определяются как факторы, которые могут ограничивать способность собирателя максимально использовать валюту. Правило оптимального решения, или лучшая стратегия поиска пищи для организма, определяется как решение, которое максимизирует валюту в условиях ограничений окружающей среды. Определение оптимального правила принятия решений - основная цель OFT.
Оптимальная модель кормодобывания генерирует количественные прогнозы того, как животные максимизируют свою приспособленность во время кормления. Процесс построения модели включает определение валюты, ограничений и соответствующего правила принятия решений для собирателя.
Валюта определяется как единица измерения, оптимизированная животным. Это также гипотеза об издержках и выгодах, которые несет это животное. Например, определенный собиратель получает энергию от пищи, но несет расходы на поиск пищи: время и энергия, потраченные на поиски, можно было бы вместо этого использовать для других целей, таких как поиск партнеров или защита детенышей. В интересах животного получить максимальную пользу при минимальных затратах. Таким образом, валюту в этой ситуации можно определить как чистый выигрыш энергии в единицу времени. Однако для другого сборщика время, необходимое для переваривания пищи после еды, может быть более значительным расходом, чем время и энергия, потраченные на поиски пищи. В этом случае валюта может быть определена как чистый прирост энергии за время оборота пищеварения вместо чистого прироста энергии за единицу времени. Кроме того, выгоды и затраты могут зависеть от сообщества собирателей. Например, собиратель, живущий в улье, скорее всего, будет добывать корм таким образом, чтобы максимизировать эффективность для его колонии, а не для него самого. Определив валюту, можно построить гипотезу о том, какие выгоды и затраты важны для рассматриваемого собирателя.
Ограничения - это гипотезы об ограничениях, налагаемых на животное. Эти ограничения могут быть связаны с особенностями окружающей среды или физиологией животного и могут ограничивать их эффективность кормления. Время, которое требуется собирателю, чтобы добраться от места гнездования до места нагула, является примером ограничения. Максимальное количество продуктов питания, которое собиратель может унести к месту своего гнездования, - еще один пример ограничения. У животных также могут быть когнитивные ограничения, такие как ограничения на обучение и память. Чем больше ограничений можно выявить в данной системе, тем большую предсказательную силу будет иметь модель.
 Рис. 1. Прирост энергии на затраты (E) для принятия стратегии кормодобывания x. Адаптировано из Parker Smith.
Рис. 1. Прирост энергии на затраты (E) для принятия стратегии кормодобывания x. Адаптировано из Parker Smith. Учитывая гипотезы о валюте и ограничениях, правило оптимального решения - это предсказание модели того, какой должна быть лучшая стратегия кормодобывания животного. Возможными примерами оптимальных правил принятия решений могут быть оптимальное количество пищевых продуктов, которые животное должно унести к месту своего гнездования, или оптимальный размер пищевых продуктов, которыми должно питаться животное. На рисунке 1 показан пример того, как оптимальное правило принятия решения может быть определено из графической модели. Кривая представляет прирост энергии на затраты (E) для принятия стратегии кормодобывания x. Прирост энергии за счет затрат - это валюта, которая оптимизируется. Ограничения системы определяют форму этой кривой. Правило оптимального решения (x *) - это стратегия, для которой валюта, выигрыш в энергии на затраты, является наибольшей. Оптимальные модели кормодобывания могут выглядеть очень по-разному и становиться очень сложными в зависимости от природы валюты и количества рассматриваемых ограничений. Однако общие принципы валюты, ограничений и правила оптимального решения остаются одинаковыми для всех моделей.
Чтобы протестировать модель, можно сравнить предсказанную стратегию с фактическим поведением животного при поиске пищи. Если модель хорошо соответствует наблюдаемым данным, то гипотезы о валюте и ограничениях поддерживаются. Если модель не соответствует данным, возможно, валюта или конкретное ограничение были неправильно определены.
Теория оптимального кормодобывания широко применяется в системах кормления во всем животном мире. Согласно OFT, любой интересующий организм можно рассматривать как хищника, добывающего добычу. Существуют разные классы хищников, в которые попадают организмы, и каждый класс имеет свои стратегии поиска пищи и хищничества.
Оптимизация этих различных стратегий поиска пищи и хищничества может быть объяснена с помощью теории оптимального кормодобывания. В каждом случае существуют затраты, выгоды и ограничения, которые в конечном итоге определяют оптимальное правило принятия решения, которому должен следовать хищник.
Одной из классических версий теории оптимального кормодобывания является модель оптимального питания, которая также известна как модель выбора добычи или модель непредвиденных обстоятельств. В этой модели хищник встречает различные предметы добычи и решает, есть ли то, что у него есть, или искать более прибыльный предмет добычи. Модель предсказывает, что собиратели должны игнорировать низкоприбыльные предметы добычи, когда более прибыльные предметы присутствуют и в изобилии.
Прибыльность предмета добычи зависит от нескольких экологических переменных. E - количество энергии (калорий), которое жертва обеспечивает хищнику. Время обработки (ч ) - это количество времени, которое требуется хищнику, чтобы обработать пищу, начиная с момента, когда хищник находит объект добычи, до момента, когда объект добычи поедается. Прибыльность добычи тогда определяется как E / h . Кроме того, время поиска (S ) - это время, которое требуется хищнику, чтобы найти объект добычи, и зависит от обилия пищи и легкости ее обнаружения. В этой модели денежной единицей является потребление энергии в единицу времени, и ограничения включают фактические значения E, hи S, а также тот факт, что объекты добычи встречаются последовательно.
Используя эти переменные, оптимальная модель диеты может предсказать, как хищники выбирают между двумя типами добычи: большая добыча 1с энергетической ценностью E1и время обработки h1, и маленькую добычу 2с энергетической ценностью E2и временем обработки h2. Чтобы максимизировать общий уровень прироста энергии, хищник должен учитывать прибыльность двух типов добычи. Если предположить, что большая добыча 1 более прибыльна, чем мелкая добыча 2, то E1/h1>E2/h2. Таким образом, если хищник сталкивается с добычей 1, он всегда должен выбирать съесть ее из-за ее более высокой прибыльности. Он никогда не должен утруждать себя поисками добычи 2. Однако, если животное встречает добычу 2, оно должно отклонить ее, чтобы искать более прибыльную добычу 1, если только не потребуется время, необходимое для поиска добычи 1 это слишком долго и дорого, чтобы того стоить. Таким образом, животное должно есть добычу 2, только если E2/h2>E1/ (h 1+S1), где S1- время поиска добычи 1. Поскольку это всегда благоприятно для выбрать съесть добычу 1, выбор съесть добычу 1 не зависит от численности добычи 2. Но поскольку длина S1(т.е. насколько сложно найти добычу1) логически зависит от плотности добычи 1, выбор съесть добычу 2 зависит от обилия добычи 1.
Модель оптимальной диеты также предсказывает, что разные типы животных должны принимать разные диеты в зависимости от различий во времени поиска. Эта идея является расширением модели выбора добычи, которая обсуждалась выше. Уравнение, E2/h2>E1/ (h 1+S1), можно переставить так, чтобы получить: S1>[(E 1h2) / E 2 ] - h 1. Эта измененная форма дает пороговое значение того, как долго S1должно быть, чтобы животное могло съесть и добычу 1, и добычу 2. Животные, у которых S1'достигают порогового значения, определяются как универсалы. В природе универсалы включают в свой рацион самые разные виды добычи. Примером универсального человека является мышь, которая потребляет большое количество семян, зерен и орехов. Напротив, хищникам с относительно короткими S1's по-прежнему лучше выбирать только добычу 1. Эти типы животных определены как специалисты и имеют очень эксклюзивный рацион по своей природе. Примером специалиста является коала, которая потребляет исключительно листья эвкалипта. В целом, разные животные, относящиеся к четырем функциональным классам хищников, демонстрируют стратегии, варьирующиеся от универсального до специализированного. Кроме того, поскольку выбор поедания жертвы 2 зависит от численности жертвы 1 (как обсуждалось ранее), если жертва 1 становится настолько редкой, что S1 достигает порогового значения, тогда животное должно переключиться с исключительно поедания добычи 1 на поедание как добычи 1, так и добычи 2. Другими словами, если в диете специалиста становится очень мало продуктов, специалист иногда может переключиться на универсального специалиста.
Как упоминалось ранее, количество времени, необходимое для поиска объекта добычи, зависит от плотности жертвы. Кривые функциональной реакции показывают скорость поимки добычи как функцию плотности пищи и могут использоваться в сочетании с теорией оптимальной диеты для прогнозирования поведения хищников при кормлении. Существует три различных типа кривых функционального ответа.
 Рисунок 2. Три типа кривых функционального ответа. Адаптировано из Staddon.
Рисунок 2. Три типа кривых функционального ответа. Адаптировано из Staddon. Для кривой функционального отклика типа I скорость захвата добычи линейно увеличивается с плотностью корма. При низкой плотности добычи время поиска велико. Поскольку хищник большую часть времени проводит в поисках, он съедает каждую находящуюся добычу. По мере увеличения плотности добычи хищник может ловить добычу все быстрее и быстрее. В определенный момент скорость поимки добычи настолько высока, что хищнику не нужно есть каждую встреченную добычу. После этой точки хищник должен выбирать только те объекты добычи с наибольшим значением E / h.
Для кривой функционального ответа Тип II скорость поимки жертвы отрицательно возрастает по мере увеличения плотность пищи. Это потому, что он предполагает, что хищник ограничен своей способностью обрабатывать пищу. Другими словами, с увеличением плотности пищи увеличивается время обработки. В начале кривой скорость поимки добычи увеличивается почти линейно с ее плотностью, и времени на обработку почти нет. По мере увеличения плотности добычи хищник тратит все меньше и меньше времени на поиск добычи и все больше и больше времени на то, чтобы обращаться с ней. Скорость поимки добычи увеличивается все меньше и меньше, пока, наконец, не выходит на плато. Большое количество добычи в основном «затопляет» хищника.
A Тип III кривая функционального ответа - это сигмовидная кривая. Скорость захвата добычи увеличивается сначала с увеличением плотности жертвы с положительным ускорением, но затем при высокой плотности изменяется на отрицательно ускоренную форму, аналогичную кривой типа II. При высокой плотности добычи (верхняя часть кривой) каждый новый объект добычи ловится практически сразу. Хищник может быть разборчивым и не ест все, что находит. Итак, предполагая, что существует два типа добычи с разной прибыльностью, оба из которых находятся в большом количестве, хищник выберет предмет с более высоким E / h . Однако при низкой плотности добычи (нижняя часть кривой) скорость поимки добычи увеличивается быстрее, чем линейно. Это означает, что по мере того, как хищник питается и тип жертвы с более высоким E / h становится менее многочисленным, хищник начинает переключать свое предпочтение на тип жертвы с более низким E / h, потому что этот тип будет относительно более многочисленным. Это явление известно как переключение добычи.
совместная эволюция хищник-жертва часто делает невыгодным для хищника поедание определенных предметов добычи, так как многие анти- защита от хищников увеличивает время обработки. Примеры включают иглы дикобраза, вкусовые качества и усвояемость ядовитой лягушки-дротика, крипсис и другие способы избегания хищников. Кроме того, поскольку токсины могут присутствовать во многих типах жертв, хищники включают в свой рацион много разнообразия, чтобы не допустить, чтобы какой-либо токсин достиг опасного уровня. Таким образом, вполне возможно, что подход, сосредоточенный только на потреблении энергии, не может полностью объяснить поведение животного в таких ситуациях при поиске пищи.
 Рисунок 3. Теорема о предельной ценности, показанная графически.
Рисунок 3. Теорема о предельной ценности, показанная графически. Теорема о предельной ценности - это тип модели оптимальности это часто применяется для оптимального кормления. Эта теорема используется для описания ситуации, в которой организм, ищущий пищу на участке, должен решить, когда ему экономически выгодно уйти. Пока животное находится внутри участка, оно испытывает закон убывающей отдачи, когда со временем становится все труднее и труднее найти добычу. Это может быть связано с тем, что добыча истощается, жертва начинает уклоняться, и ее становится труднее поймать, или хищник начинает чаще пересекать свой собственный путь в процессе поиска. Этот закон убывающей отдачи можно представить как кривую увеличения энергии за время, проведенное в заплате (рис. 3). Кривая начинается с крутого наклона и постепенно выравнивается по мере того, как добычу становится труднее найти. Еще одна важная стоимость, которую следует учитывать, - это время в пути между разными участками и местом гнездования. Животное теряет время на поиск пищи во время путешествия и расходует энергию на свое движение.
В этой модели оптимизируемой валютой обычно является чистый прирост энергии за единицу времени. Ограничениями являются время прохождения и форма кривой убывающей отдачи. Графически валюта (чистый прирост энергии в единицу времени) задается наклоном диагональной линии, которая начинается в начале времени путешествия и пересекает кривую убывающей доходности (Рисунок 3). Чтобы максимизировать валюту, нужна линия с наибольшим наклоном, которая все еще касается кривой (касательная линия). Место, где эта линия касается кривой, обеспечивает оптимальное правило принятия решения относительно количества времени, которое животное должно провести на участке перед тем, как уйти.
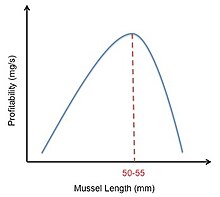 Рис. 4. Кривая прибыльности мидий, сдвинутая вправо. По материалам Meire Ervynck.
Рис. 4. Кривая прибыльности мидий, сдвинутая вправо. По материалам Meire Ervynck. Охотник-сорока кормление мидиями является примером того, как можно использовать оптимальную модель рациона. Ловцы-сороки кормятся мидиями и раскалывают их клювами. Ограничения для этих птиц - характеристики мидий разного размера. В то время как большие мидии дают больше энергии, чем маленькие, большие мидии сложнее вскрыть из-за их более толстой оболочки. Это означает, что хотя крупные мидии имеют более высокое содержание энергии (E ), они также имеют более длительное время обработки (ч ). Доходность любой мидии рассчитывается как E / h . Ловцы-сороки должны решить, какой размер мидии обеспечит достаточное питание, чтобы перевесить затраты и энергию, необходимые для ее открытия. В своем исследовании Мейре и Эрвинк попытались смоделировать это решение, построив график относительной прибыльности мидий разного размера. Они получили колоколообразную кривую, указывающую на то, что мидии среднего размера были наиболее прибыльными. Однако они заметили, что, если ловец-сорока отказывался от слишком большого количества мелких мидий, время, необходимое для поиска следующей подходящей мидии, значительно увеличивалось. Это наблюдение сместило их колоколообразную кривую вправо (рис. 4). Однако, хотя эта модель предсказывала, что кулики-сороки должны отдавать предпочтение мидиям размером 50–55 мм, полученные данные показали, что на самом деле ловцы-сороки предпочитают мидий размером 30–45 мм. Затем Мейре и Эрвинк поняли, что предпочтение размера мидий зависит не только от прибыльности добычи, но и от плотности добычи. После того, как это было учтено, они обнаружили хорошее соответствие между предсказанием модели и наблюдаемыми данными.
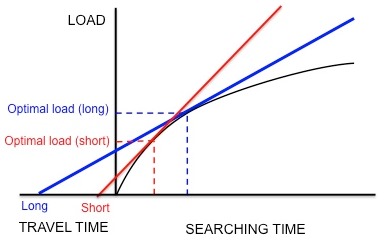 Рис. 5. Если скворцы максимизируют чистую скорость прироста энергии, увеличивается время путешествия. при большей оптимальной нагрузке. По материалам Кребса и Дэвиса.
Рис. 5. Если скворцы максимизируют чистую скорость прироста энергии, увеличивается время путешествия. при большей оптимальной нагрузке. По материалам Кребса и Дэвиса. Кормление европейского скворца, Sturnus vulgaris, является примером того, как теорема о предельной ценности используется для моделирования оптимального кормления. Скворцы покидают свои гнезда и отправляются на пищевые участки в поисках личинок кожаных курток, чтобы вернуть своих детенышей. Скворцы должны определить оптимальное количество предметов добычи, которые они могут забрать за одну поездку (то есть оптимальный размер груза). Пока скворцы добывают корм в пределах грядки, отдача от них уменьшается: скворец может удерживать в своем клюве лишь определенное количество кожаных курток, поэтому скорость, с которой родитель захватывает личинок, уменьшается вместе с количеством личинок, уже имеющихся в клюве.. Таким образом, ограничениями являются форма кривой убывающей отдачи и время прохождения (время, необходимое для того, чтобы совершить путешествие туда и обратно от гнезда до участка и обратно). Кроме того, предполагается, что валюта представляет собой чистый выигрыш энергии в единицу времени. Используя эту валюту и ограничения, можно спрогнозировать оптимальную нагрузку, проведя касательную линию к кривой убывающей доходности, как обсуждалось ранее (рисунок 3).
Касельник и др. хотел определить, действительно ли этот вид оптимизирует чистый прирост энергии в единицу времени, как предполагалось. Они разработали эксперимент, в котором скворцов обучали собирать мучных червей из искусственной кормушки на разном расстоянии от гнезда. Исследователи искусственно создали фиксированную кривую убывающей отдачи для птиц, сбрасывая мучных червей через все более длительные интервалы. Птицы продолжали собирать мучных червей по мере их предъявления, пока они не достигли «оптимальной» нагрузки и не улетели домой. Как показано на рисунке 5, если бы скворцы максимизировали чистый прирост энергии в единицу времени, короткое время в пути предсказывало бы небольшую оптимальную нагрузку, а большое время в пути предсказывало бы большую оптимальную нагрузку. В соответствии с этими прогнозами Качельник обнаружил, что чем больше расстояние между гнездом и искусственной кормушкой, тем больше размер груза. Кроме того, наблюдаемые размеры нагрузки количественно очень точно соответствовали прогнозам модели. Другие модели, основанные на разных валютах, таких как количество энергии, полученной за счет затраченной энергии (т.е. энергоэффективность), не смогли точно предсказать наблюдаемые размеры нагрузки. Таким образом, Качельник пришел к выводу, что скворцы максимизируют чистый прирост энергии в единицу времени. Этот вывод не был опровергнут в более поздних экспериментах.
Рабочие пчелы представляют собой еще один пример использования теоремы о предельной ценности при моделировании оптимального кормового поведения. Пчелы переходят от цветка к цветку, собирая нектар, чтобы отнести его обратно в улей. Хотя эта ситуация похожа на ситуацию со скворцами, и ограничения, и валюта на самом деле у пчел разные.
Пчела не получает убывающей урожайности из-за истощения нектара или каких-либо других характеристик самих цветов. Общее количество собранного нектара линейно увеличивается со временем, проведенным на грядке. Однако вес нектара значительно увеличивает стоимость полета пчелы между цветами и ее обратного пути в улей. Вольф и Шмид-Хемпель показали, экспериментируя с разными грузами на спинах пчел, что стоимость тяжелого нектара настолько высока, что пчелы сокращают продолжительность жизни. Чем короче продолжительность жизни рабочей пчелы, тем меньше времени у нее остается на жизнь своей колонии. Таким образом, существует кривая убывающей отдачи от чистого выхода энергии, которую получает улей по мере того, как пчела собирает больше нектара за одну поездку.
Стоимость тяжелого нектара также влияет на валюту, используемую пчелами. В отличие от скворцов в предыдущем примере, пчелы максимизируют энергоэффективность (количество энергии, полученной на затраченную энергию), а не чистую скорость прироста энергии (чистую энергию, полученную за время). Это связано с тем, что оптимальная нагрузка, прогнозируемая за счет максимального увеличения чистого прироста энергии, слишком велика для пчел и сокращает их продолжительность жизни, снижая их общую продуктивность для улья, как объяснялось ранее. Максимально повышая энергоэффективность, пчелы могут не тратить слишком много энергии на поездку и могут прожить достаточно долго, чтобы максимально увеличить продуктивность своего улья за всю жизнь. В другой статье Шмид-Хемпель показал, что наблюдаемая взаимосвязь между размером груза и временем полета хорошо коррелирует с прогнозами, основанными на максимизации энергоэффективности, но очень плохо коррелирует с прогнозами, основанными на максимальном чистом приросте энергии.
Природа выбора добычи двумя центрархидами (белым краппи и синежабриками) была представлена Манатунджем и Асаэдой как модель, включающая оптимальные стратегии поиска пищи. Поле зрения кормящейся рыбы, представленное реактивным расстоянием, было подробно проанализировано для оценки количества встреч с добычей за один поисковый бой. Расчетные реакционные расстояния сравнивались с экспериментальными данными. Энергетические затраты, связанные с поведением рыб при поиске пищи, рассчитывались на основе последовательности событий, которые имеют место для каждой съеденной добычи. Сравнение относительной численности видов и категорий размеров добычи в желудке с окружающей средой озера показало, что и белый краппи, и синежабер (длина < 100 mm) strongly select prey utilizing an energy optimization strategy. In most cases, the fish exclusively selected large Daphnia ignoring evasive prey types (Cyclops, Diaptomids) and small cladocera. This selectivity is the result of fish actively avoiding prey with high evasion capabilities even though they appear to be high in energetic content and having translated this into optimal selectivity through capture success rates. The energy consideration and visual system, apart from the forager's ability to capture prey, are the major determinants of prey selectivity for large-sized bluegill and white crappie still at planktivorous stages.
Хотя многие исследования, такие как приведенные в примерах выше, обеспечивают количественную поддержку теории оптимального кормодобывания и демонстрируют ее полезность, модель подверглась критике за ее валидность и ограничения.
Во-первых, теория оптимального кормодобывания основывается на предположении, что естественный отбор оптимизирует стратегии добычи пищи организмов. Однако естественный отбор - это не всемогущая сила, которая производит совершенные замыслы, а скорее пассивный процесс отбора по генетически обусловленным признакам, которые увеличивают репродуктивный успех организмов . Учитывая, что генетика включает взаимодействия между локусами, рекомбинацией и другие сложности, нет гарантии, что естественный отбор может оптимизировать специфический поведенческий параметр.
Кроме того, OFT также предполагает, что пищевое поведение может свободно формироваться естественным отбором, поскольку это поведение не зависит от других видов деятельности организма. Однако, учитывая, что организмы представляют собой интегрированные системы, а не механические агрегаты частей, это не всегда так. Например, необходимость избегать хищников может заставить собирателей кормиться меньше оптимальной нормы. Таким образом, пищевое поведение организма не может быть оптимизировано, как можно было бы прогнозировать с помощью OFT, потому что оно не является независимым от другого поведения.
Еще одно ограничение OFT состоит в том, что ему не хватает точности на практике. Теоретически оптимальная модель кормодобывания дает исследователям конкретные количественные прогнозы об оптимальном правиле принятия решений хищником, основанном на гипотезах о валюте и ограничениях системы. Однако на самом деле трудно определить основные понятия, такие как тип добычи, частота встреч или даже участок, как их воспринимает собиратель. Таким образом, хотя переменные OFT теоретически могут казаться согласованными, на практике они могут быть произвольными и трудными для измерения.
Кроме того, хотя предпосылка OFT состоит в том, чтобы максимизировать приспособленность организма, многие исследования показывают только корреляцию между наблюдаемым и прогнозируемым поведением при поиске пищи и не проверяют, действительно ли поведение животного увеличивает его репродуктивную пригодность. Возможно, что в некоторых случаях корреляция между отдачей от добычи пищи и репродуктивным успехом вообще отсутствует. Без учета этой возможности многие исследования, использующие OFT, остаются неполными и не рассматривают и не проверяют основную идею теории.
Одна из самых настоятельных критических замечаний в отношении OFT состоит в том, что его нельзя по-настоящему проверить. Эта проблема возникает всякий раз, когда есть расхождения между прогнозами модели и фактическими наблюдениями. Трудно сказать, ошибочна ли модель в корне, или конкретная переменная была идентифицирована неточно или не учтена. Поскольку в модель можно добавлять бесконечные правдоподобные модификации, модель оптимальности никогда не может быть отвергнута. Это создает проблему, заключающуюся в том, что исследователи формируют свою модель в соответствии со своими наблюдениями, вместо того, чтобы тщательно проверять свои гипотезы о поведении животных в поисках пищи.