
Войти
O-GlcNAc (сокращение от O-связанный GlcNAc или O-связанный β-N-ацетилглюкозамин ) является обратимым ферментная посттрансляционная модификация, которая обнаруживается на сериновых и треониновых остатках нуклеоцитоплазматических белков. Модификация представляет β-гликозидной связью между гидроксильной группой сериновых или треониновых боковых цепей и N-ацетилглюкозамином (GlcNAc). O-GlcNAc от других форм гликозилирования белка: (i) O-GlcNAc не удлиняется и не модифицируется с образованием более сложных структур гликанов, (ii) O-GlcNAc почти исключительно обнаружен на ядерных и цитоплазматических белках, а не на мембранных белках и секреторных белках, и (iii) O-GlcNAc представляет собой высокодинамичную модификацию, которая изменяется быстрее, чем белки, которые она модифицирует. O-GlcNAc консервативен в серине многоклеточных животных.
 в полипептиде без модификаций (вверху) и с модификацией O-GlcNAc (внизу). (PDB : 4GYW)
в полипептиде без модификаций (вверху) и с модификацией O-GlcNAc (внизу). (PDB : 4GYW) Из-за динамической природы O-GlcNAc и его присутствие на остатках серина и треонина, O-GlcNAцилирование в некоторых отношениях сходно с фосфорилированием белка. Хотя существует примерно 500 киназ и 150 фосфатаз, которые регулируют фосфорилирование белков у человека, есть только 2 фермента, которые регулируют цикл O-GlcNAc: трансфераза O-GlcNAc (OGT) и O-GlcNAcase (OGA) катализируют добавление и удаление O-GlcNAc соответственно. OGT использует UDP-GlcNAc в качестве сахара-донора для переноса сахара.
Впервые описанная в 1984 году, посттрансляционная модификация с тех пор идентифицирована для более чем 4000 белков. Сообщалось о множестве функциональных ролей для O-GlcNAcylation, включая перекрестное взаимодействие с фосфорилированием серина / треонина, регулирование белок-белковых взаимодействий, изменение структуры белка или активность фермента, изменение субклеточной локализации белка , и модулирование стабильности и деградации белка. Было обнаружено, что многие компоненты клеточного аппарата транскрипции модифицируют O-GlcNAc, и во многих исследованиях сообщается о связи между O-GlcNAc, транскрипцией и эпигенетикой. O-GlcNAc влияет на многие другие клеточные процессы, такие как апоптоз, клеточный цикл и стрессовые реакции. Средство UDP-GlcNAc является конечным продуктом пути биосинтеза гексозамина, который объединяет метаболу, углевод, жирную кислоту и нуклеотид метаболизма, было предположение, что O-GlcNAc действует как «датчик питательных веществ » и реагирует на метаболический статус клетки. Нарушение регуляции O-GlcNAc вовлечено во многие патологии, включая болезнь Альцгеймера, рак, диабет и нейродегенеративные расстройства.
 GalT-радиоактивное мечение клеточных белков с помощью UDP- [H] галактоза с последующим β-элиминированием давала Galβ1-4GlcNAcitol, что позволяет предположить, что субстратом для GalT был O-GlcNAc. Радиоактивно меченая [H] галактоза первого красным.
GalT-радиоактивное мечение клеточных белков с помощью UDP- [H] галактоза с последующим β-элиминированием давала Galβ1-4GlcNAcitol, что позволяет предположить, что субстратом для GalT был O-GlcNAc. Радиоактивно меченая [H] галактоза первого красным. В 1984 г. лаборатория Hart исследовала концевые остатки GlcNAc на поверхности тимоцитов и лимфоцитов. β-1,4-галактозилтрансфераза коровьего молока, которая реагирует с концевыми остатками GlcNAc, была для проведения радиоактивного мечения UDP- [H] галактозой. β-элиминация к белкам O-гликозидно; хроматография показала, что основным продуктом β-элиминирования был Galβ1-4GlcNAcitol. Нечувствительность к лечению пептидной N-гликозидазой предоставила дополнительные доказательства наличия O-компонентов GlcNAc. Повышение проницаемости клеток детергентом перед радиоактивной меткой значительно увеличило количество [H] галактозы, включенной в Galβ1-4GlcNAcitol, что привело к выводу, что большая часть связанных моносахаридных остатков GlcNAc находится внутриклеточно.
 O-GlcNAcилирование остатков серина и треонина динамически контролируется OGT и OGA.
O-GlcNAcилирование остатков серина и треонина динамически контролируется OGT и OGA. O-GlcNAc обычно представляет собой динамическую модификацию, которая может циклически включать и выключать белки. Считается, что некоторые остатки конститутивно модифицируются O-GlcNAc. Модификация O-GlcNAc устанавливает OGT в последовательном би-би-механизме, где сахар-донор, UDP-GlcNAc, сначала связывается с OGT, а с белком-субстратом. Модификация O-GlcNAc удаляется с помощью OGA в механизме гидролиза, включающего анхимерную помощь (катализ с помощью субстрата), чтобы получить немодифицированный белок и GlcNAc. Хотя кристаллические структуры демонстрируют как для OGT, так и для OGA, точные механизмы, с помощью OGT и OGA распознают субстраты, полностью не установлены. В отличие от N-связанного гликозилирования, для которого гликозилирование происходит в конкретной консенсусной последовательности (Asn-X-Ser / Thr, где X - любая аминокислота, кроме Pro), нет окончательной консенсусной последовательность был идентифицирован для O-GlcNAc,. Следовательно, прогнозирование сайтов модификации O-GlcNAc является сложной процедурой, и для определения сайтов модификации обычно требуются методы масс-спектрометрии. Для OGT исследования показали, что распознавание субстрата регулируется рядом факторов, включая лестничные мотивы аспартата и аспарагина в просвете супспирального домена TPR, активные остатки сайта и адаптерные белки. Предположительно, что OGT требует, чтобы его структура была в расширенной конформации, что OGT отдает предпочтение гибким подложкам. В кинетических экспериментах in vitro, измеряющих активность OGT и OGA на панели белковых субстратов, было показано, что кинетические параметры OGT, различные между различными белками, в то время как кинетические параметры для OGA были относительно постоянными для разных белков. Этот результат свидетельствует о том, что OGT является «старшим партнером» в регуляции O-GlcNAc, а OGA в первую распознает субстраты через присутствие O-GlcNAc, а не идентичность модифицированного белка.
 Слева: Модель полноразмерного ncOGT в комплексе с пептидным субстратом CKII и UDP. Цвета указывает на домен TPR (серый), N-концевую область каталитического домена (светло-розовый), промежуточный домен (светло-зеленый), C-концевую область каталитического домена (голубой), пептидный субстрат CKII (зеленый) и UDP (голубой).). Справа: структура димера OGA D175N человека в комплексе с O-GlcNAцилированным пептидным субстратом TAB1. Мономеры показаны сине-белым / светло-желтым, соответствующие пептидные субстраты - синим / желтым. (PDB: 5VVU)
Слева: Модель полноразмерного ncOGT в комплексе с пептидным субстратом CKII и UDP. Цвета указывает на домен TPR (серый), N-концевую область каталитического домена (светло-розовый), промежуточный домен (светло-зеленый), C-концевую область каталитического домена (голубой), пептидный субстрат CKII (зеленый) и UDP (голубой).). Справа: структура димера OGA D175N человека в комплексе с O-GlcNAцилированным пептидным субстратом TAB1. Мономеры показаны сине-белым / светло-желтым, соответствующие пептидные субстраты - синим / желтым. (PDB: 5VVU) Существует несколько методов обнаружения присутствия O-GlcNAc и характеристик модифицированных остатков.
Агглютинин зародышей пшеницы, растительный лектин, способен распознавать концевые остатки GlcNAc и поэтому часто используется для обнаружения O-GlcNAc. Этот лектин применен в лектиновой аффинной хроматографии для обогащения и обнаружения O-GlcNAc.
Pan-O-GlcNAc антитела которые распознают модификацию O-GlcNAc в степени независимо от идентичности модифицированного белка. К ним против RL2, антитело IgG, индуцированное против O-GlcNAцилированных белков комплекса ядерных пор, и CTD110.6, антитело IgM, индуцированное против иммуногенного пептида с помощью одинарная сериновая модификация O-GlcNAc. Сообщалось о других O-GlcNAc-специфических антителах, которые имели некоторую зависимость от идентичности модифицированного белка.
Многие метаболические химические репортеры разработаны для идентификации O-GlcNAc. Метаболические химические репортеры, как правило, имеют аналоги сахаров, которые несут дополнительную химическую составляющую, обеспечивающую дополнительную реактивность. Например, перацетилированный GlcNAc (Ac 4 GlcNAz) представляет собой проницаемый для клеток азидо сахар, который деэтерифицируется внутриклеточно эстеразами в GlcNAz и превращается в UDP- GlcNAz в пути спасения гексозамина. UDP-GlcNAz может установить OGT в качестве донора сахара для введения модификации O-GlcNAz. Присутствие азидосахара затем можно визуализировать с помощью алкин -содержащих биоортогональных химических зондов в реакции азид-алкинового циклоприсоединения. Эти зонды могут идентифицироваться легко идентифицируемые метки, такие как пептид FLAG, биотин и молекулы красителя. Массовые метки на основе полиэтиленгликоля (PEG) также использовались для измерения стехиометрии O-GlcNAc. Конъюгация молекул ПЭГ 5 кДа приводит к массовому получению белков - белков с более высокой степенью O-GlcNAцилированных белков будут несколько иметь молекулы ПЭГ и таким образом, мигрировать медленнее при гель-электрофорезе. Сообщалось о других химических химических репортерах, несущих азиды или алкины (обычно в положениях 2 или 6). Вместо Gl аналоговcNAc можно использовать аналоги GalNAc, а также UDP-GalNAc находится в равновесии с UDP-GlcNAc в клетках из-за действия UDP-галактоза-4'-эпимеразы (GALE). Было обнаружено, что обработка Ac 4 GalNAz приводит к усилению мечения O-GlcNAc по сравнению с Ac 4 GlcNAz, возможно, из-за узкого места в UDP-GlcNAc пирофосфорилазе обработка GlcNAz-1-P в UDP-GlcNAz. Ac 3 GlcN-β-Ala-NBD-α-1-P (Ac-SATE) 2, метаболический химический репортер, который внутриклеточно обрабатывается до меченного флуорофором UDP-GlcNAc аналог, как было показано, обеспечивает одностадийное флуоресцентное мечение O-GlcNAc в живых клетках.
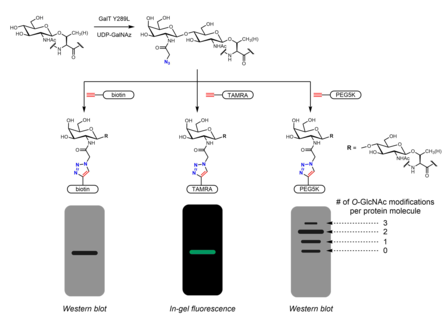 Химиоферментное мечение для обнаружения O-GlcNAc. GalT Y289L передает GalNAz в O-GlcNAc, механизм для химии щелчков. Различные зонды можно конъюгировать посредством азид-алкинового циклоприсоединения. Присоединение масс-метки PEG5K позволяет визуализировать стехиометрию O-GlcNAc.
Химиоферментное мечение для обнаружения O-GlcNAc. GalT Y289L передает GalNAz в O-GlcNAc, механизм для химии щелчков. Различные зонды можно конъюгировать посредством азид-алкинового циклоприсоединения. Присоединение масс-метки PEG5K позволяет визуализировать стехиометрию O-GlcNAc. Метаболическое мечение также может быть для обозначения партнеров по связыванию O-GlcNAцилированных белков. N-ацетильная группа может быть удлиненной для включения фрагмента диазирина. Обработка клеток перацетилированным фосфат-защищенным Ac 3 GlcNDAz-1-P (Ac-SATE) 2 приводит к модификации белков с помощью O-GlcNDAz. Затем УФ-облучение индуцирует фотосшивание между белками, несущими модификацию O-GlcNDaz, взаимодействующими белками.
Были выявлены некоторые проблемы с различными химическими химическими репортерами, например, их использование может ингибировать путь биосинтеза гексозамина, они не распознаются с помощью OGA и, следовательно, не способны улавливать цикл O-GlcNAc, или они могут быть включены в модификации гликозилирования дополнительных O-GlcNAc, как это видно в секретируемых белках. Метаболические химические репортеры с химическими маркерами в N-ацетильном положении могут также маркировать ацетилированные белки, поскольку ацетильная группа может гидролизоваться до ацетатных аналогов, которые могут использоваться для ацетилирования белков.
Хемоэнзиматическая маркировка альтернативной стратегию включения ручек для химии щелчков. Система ферментной маркировки Click-IT O-GlcNAc, разработанная группа Hsieh-Wilson и коммерциализированная Invitrogen, использует мутантный фермент GalT Y289L, способный переносить азидогалактозу (GalNAz) на O-GlcNAc. Присутствие GalNAz (и, следовательно, также O-GlcNAc) может быть обнаружено с помощью различных алкинсодержащих зондов, идентифицируемых метками, такими как биотин, молекулы красителя и PEG.
 биосенсор FRET для O-GlcNAc. В условиях высокого содержания O-GlcNAc GafD будет связывать группу O-GlcNAc на пептидном субстрате CKII, сближая CFP и YFP для FRET. Различные системы локализации могут быть объединены, чтобы локализовать датчик в различных клеточных компартментах, например ядре, цитоплазме и плазматической мембране.
биосенсор FRET для O-GlcNAc. В условиях высокого содержания O-GlcNAc GafD будет связывать группу O-GlcNAc на пептидном субстрате CKII, сближая CFP и YFP для FRET. Различные системы локализации могут быть объединены, чтобы локализовать датчик в различных клеточных компартментах, например ядре, цитоплазме и плазматической мембране. Разработан белковый биосенсор, который может обнаруживать изменения в уровнях O-GlcNAc с использованием резонансной передачи энергии Фёрстера. Этот сенсор состоит из четырех компонентов, связанных вместе в следующем порядке: голубой флуоресцентный белок (CFP), связывающий домен O-GlcNAc (на основе GafD, лектина, чувствительность к концевому β - O -GlcNAc), пептид CKII, который является известным субстратом OGT, и желтый флуоресцентный белок (YFP). После O-GlcNA-цилирования пептида CKII домен GafD связывает фрагмент O-GlcNAc, сближая домены CFP и YFP и генерируя сигнал FRET. Генерация этого сигнала обратима и другая группа для мониторинга динамики O-GlcNAc в ответ на различные виды лечения. Этот датчик может быть генетически закодирован и познакомиться в клетках. Добавление последовательности локализации позволяет нацеливать этот датчик O-GlcNAc на ядро, цитоплазму или плазматическую мембрану.
Биохимические подходы, такие как Вестерн-блоттинг может предоставить подтверждающие доказательства того, что белок модифицирован O-GlcNAc; масс-спектрометрия (МС) может предоставить окончательные доказательства присутствия O-GlcNAc. Гопротеомные исследования с применением МС способствовали идентификации белков модифицированных O-GlcNAc.
Буква O-GlcNAc является субстехиометрическим и ионным подавлением происходит в присутствии немодифицированных пептидов, стадию обогащения обычно проводят перед масс-спектрометрическим анализом. Это можно сделать с помощью лектинов, антител или химической маркировки. Модификация O-GlcNAc лабильна в отношении методов фрагментации, индуцированной столкновения, таких как диссоциация, индуцированная столкновение (CID) и столкновение этих методов с более высокой энергией (HCD), поэтому эти методы изолированы не всегда применимы для картирования сайтов O-GlcNAc. HCD генерирует фрагментные ионы, характерные для N-ацетилгексозаминов, которые можно использовать для определения статуса O-GlcNAцилирования. Для облегчения картирования сайта с помощью HCD для преобразования лабильной модификации O-GlcNAc в более стабильную массовую метку можно использовать β-элиминацию с последующим добавлением Майкла с дитиотреитолом (BEMAD).. Для BEMAD-картирования O-GlcNAc необходимо обработать фосфататазой, иначе могут быть обнаружены другие посттрансляционные модификации серина / треонина, такие как фосфорилирование. Диссоциация с переносом электрона (ETD) используется для картирования сайта, как ETD вызывает расщепление пептидного остова, оставляя посттрансляционные модификации, такие как O-GlcNAc, нетронутыми.
Традиционные протеомные исследования проводят тандемный MS на наиболее распространенных масс-спектрах полного сканирования, что запрещает полную характеристику с низкой численностью. Одна современная стратегия направленной протеомики использует изотопные метки, например дибромид, для маркировки O-GlcNAцилированных белков. Этот метод позволяет алгоритмически обнаруживать виды с низкой численностью, которые затем секвенируются тандемной МС. Направленная тандемная МС и целевое назначение гликопептидов идентифицировать последовательность O-GlcNAцилированного пептида. Один пример зонда состоит из биотиновой аффинной метки, расщепляемого кислотой силана, мотива перекодирования изотопов и алкина. Однозначное картирование сайта возможно для пептидов только с одним остатком серин / треонин.
Общая процедура для этого метода гликопротеомики, нацеливания на изотопы (IsoTaG), следующая:
 Структура зонда IsoTaG. Зонд состоит из биотиновой аффинной метки (красный), линкера (черный), расщепляемого кислотой силана (синий), мотива перекодирования изотопа (зеленый) и алкина (фиолетовый).
Структура зонда IsoTaG. Зонд состоит из биотиновой аффинной метки (красный), линкера (черный), расщепляемого кислотой силана (синий), мотива перекодирования изотопа (зеленый) и алкина (фиолетовый). Для количественного профилирования O-GlcNAc с использованием дифференциального изотопного мечения. Примеры зондов обычно состоят из аффинной метки биотина, расщепляемого линкера (кислотного или фоторасщепляемого), тяжелой или легкой изотопной метки и алкина.
Были разработаны различные химические и генетические стратегии для манипулирования O-GlcNAc как на основе протеома, так и на конкретных белках.
Сообщалось о низкомолекулярных ингибиторах как для OGT, так и для OGA, которые действуют в клетках или in vivo. Ингибиторы OGT приводят к глобальному снижению O-GlcNAc, тогда как ингибиторы OGA приводят к глобальному увеличению O-GlcNAc; эти ингибиторы не способны модулировать O-GlcNAc на определенных белках.
Ингибирование пути биосинтеза гексозамина также способно снизить уровни O-GlcNAc. Например, аналоги глутамина азасерин и 6-диазо-5-оксо-L-норлейцин (DON) могут ингибировать GFAT, хотя эти молекулы также могут не- специфически влияют на другие пути.
Лигирование экспрессированного белка использовалось для получения белков, модифицированных O-GlcNAc, сайт-специфическим образом. Существуют методы твердофазного пептидного синтеза с включением GlcNAc-модифицированного серина, треонина или цистеина.
 Сайт-направленный мутагенез для манипулирования O-GlcNAc. Мутагенез S / T-t o-A предотвращает модификацию O-GlcNAc по этому остатку. Мутагенез S / T-to-C позволяет генерировать модификацию S-GlcNAc, структурный аналог O-GlcNAc, который нелегко гидролизуется OGA.
Сайт-направленный мутагенез для манипулирования O-GlcNAc. Мутагенез S / T-t o-A предотвращает модификацию O-GlcNAc по этому остатку. Мутагенез S / T-to-C позволяет генерировать модификацию S-GlcNAc, структурный аналог O-GlcNAc, который нелегко гидролизуется OGA. Сайт-направленный мутагенез остатков серина или треонина модифицированных O-GlcNAc к аланину можно использовать для оценки функций O-GlcNAc по конкретным остаткам. Таким образом, боковая цепь аланина представляет собой метильную группу и таким образом, не может действовать как сайт O-GlcNAc, эта мутация эффективно навсегда удаляет O-GlcNAc по определенному остатку. Хотя фосфорилирование серина / треонина можно смоделировать с помощью мутагенеза до аспартата или глутамата, которые имеют отрицательно заряженные боковые цепи карбоксилата, ни одна из 20 канонических аминокислот не воспроизводит в достаточной степени свойства свойства O-GlcNAc. Мутагенез до триптофана был использован для имитации стерической массы O-GlcNAc, хотя триптофан намного более гидрофобен, чем O-GlcNAc. Мутагенез может также нарушать другие посттрансляционные модификации, например, если серин альтернативно фосфорилирован или O-GlcNAцилирован, мутагенез аланина навсегда устраняет возможности как фосфорилирования, так и O-GlcNAцилирования.
Масс-спектрометрия идентифицировала S-GlcNAc как посттрансляционную модификацию, обнаруженную на остатках цистеина. Эксперименты in vitro применяли, что OGT может катализировать образование S-GlcNAc и что OGA не может гидролизовать S-GlcNAc. Хотя в предыдущем сообщении предполагалось, что OGA способен гидролиз тиогликозиды, это было сделано только на арилтиогликозид пара-нитрофенол-S-GlcNAc; пара-нитротиофенол является более активированной уходящей группой, чем остаток цистеина. Недавние исследования подтвердили использование S-GlcNAc в качестве ферментативно стабильной структурной модели O-GlcNAc, которая может быть включена посредством твердофазного пептидного синтеза или сайта-направленного мутагенеза.
Слитые конструкции из нанотела и OGT с усеченным TPR допускают индуцированное близостью специфичное для белка O-GlcNAцилирование в клетках. Нанотело может быть направлено на белковые метки, например, GFP, которые слиты с целевым белком, или нанотело могут быть направлены на эндогенные белки. Например, нанотело, распознающее С-концевую последовательность EPEA, может направлять ферментативную активность OGT на α-синуклеин.
Апоптоз, форма управляемой гибель клеток, как решено, регулируется O-GlcNAc. Сообщалось, что при различных раковых заболеваниях повышенные уровни O-GlcNAc подавляют апоптоз. Сообщалось, что каспаза-3, каспаза-8 и каспаза-9 быть модифицированным O-GlcNAc. Каспаза-8 модифицируется вблизи сайтов расщепления / активации; Модификация O-GlcNAc может блокировать расщепление и активацию каспазы-8 из-за стерических затруднений. Фармакологическое снижение O-GlcNAc с помощью 5S-GlcNAc ускоряет активацию каспазы, в то время как фармакологическое повышение O-GlcNAc с помощью тиамет-G ингибирует активацию каспазы.
Белки, которые регулируют генетику, часто делятся на категории писателей, читателей и стиральных машин, то есть ферменты, которые устанавливают эпигенетические модификации, белки, распознающие эти модификации, и ферменты, которые удаляют эти модификации. На сегодняшний день O-GlcNAc идентифицирован на фермах писателя и ластика. O-GlcNAc обнаруживается во многих местах на EZH2, каталитической метилтрансферазе субъединице PRC2, и считается, что он стабилизирует EZH2 до образования комплекса PRC2 и регулирует ди - и активность триметилтрансферазы. Все три члена семейства транслокаций десять-одиннадцать (TET) диоксигеназ (TET1, TET2 и TET3 ) известны как модифицированные O-GlcNAc. Было высказано предположение, что O-GlcNAc вызывает ядерный экспорт TET3, снижая его ферментативную активность, истощая его из ядра. O-GlcNAцилирование HDAC1 связано с повышенным активирующим фосфорилированием HDAC1.
Гистон белки, основной белковый компонент хроматина, как известно, модифицированные O-GlcNAc. O-GlcNAc был идентифицирован на всех гистонах ядра (H2A, H2B,H3 и H4 ). Было высказано предположение, что присутствие O-GlcNAc на гистонах влияет на транскрипцию гена, а также на другие гистоновые метки, такие как ацетилирование и моноубиквитинирование. Сообщалось, что TET2 взаимодействует с доменом TPR OGT и способствуют привлечению OGT к гистонам. Это взаимодействие связано с H2B S112 O-GlcNAc, который, в свою очередь, связан с моноубиквитинированием H2B K120. Было обнаружено, что фосфилирование OGT T444 через AMPK ингибирует ассоциацию OGT-хроматина и подавляет регуляцию H2B S112 O-GlcNAc.
Продукт пути биосинтеза гексозамина, UDP -GlcNAc используется OGT для катализирования добавления O-GlcNAc. Этот путь объединяет информацию о системе обмена веществ, включая аминокислоты, углеводы, жирные кислоты и нуклеотиды. Следовательно, уровни UDP-GlcNAc чувствительны к уровню клеточных метаболитов. Активность OGT частично регулируется концентрацией UDP-GlcNAc, что создает связь между статусом питательных веществ клетки и O-GlcNAc.
Депривация глюкозы вызывает снижение уровней UDP-GlcNAc и начальное снижение O-GlcNAc, но как ни странно, O-GlcNAc позже значительно активируется. Было показано, что это более активное увеличение зависит от активации AMPK и p38 MAPK, и эффект частично обусловлен повышением уровней мРНК OGT и белка. Было также высказано предположение, что этот эффект зависит от кальция и CaMKII. Активированный p38 способен рекрутировать OGT на другие белковые мишени, включая нейрофиламент H ; Модификация нейрофиламента H O-GlcNAc увеличивает его растворимость. Во время депривации глюкозы гликогенсинтаза модифицируется O-GlcNAc, который ингибирует его активность.
NRF2, фактор транскрипции, связанный с Было обнаружено, что клеточный ответ на окислительный стресс косвенно регулируется O-GlcNAc. KEAP1, адаптерный белок для cullin 3 -зависимого комплекса E3 убиквитин-лигаза, опосредует деградацию NRF2; окислительный стресс приводит к конформационным изменениям в KEAP1, которые подавляют деградацию NRF2. Модификация O-GlcNAc KEAP1 на S104 необходима для эффективного убиквитинирования и деградации NRF2, связывая O-GlcNAc с окислительным стрессом. Недостаток глюкозы приводит к снижению O-GlcNAc и снижает деградацию NRF2. Клетки, экспрессирующие мутант KEAP1 S104A, устойчивый к индуцированному ластином ферроптозу, что соответствует более высокому уровню NRF2 при удалении S104 O-GlcNAc.
Повышенные уровни O-GlcNAc были связаны с уменьшением синтеза печеночного глутатиона, важного клеточного антиоксиданта. Передозировка ацетаминофена приводит к накоплению в печени сильно окисляющего метаболита NAPQI, который детоксицируется глутатионом. У мышей нокаут OGT оказывает защитное действие против повреждений, вызванных ацетаминофеном, в то время как ингибирование OGA с помощью тиамет-G усугубляет повреждение печени, вызванное ацетаминофеном.
Обнаружено для замедления агрегации белков, хотя общность этого явления неизвестна.
Твердофазный пептидный синтез использовали для получения полноразмерного α-синуклеина с модификацией O-GlcNAc на T72. Анализы агрегации тиофлавина T и просвечивающая электронная микроскопия соответствует, что этот модифицированный α-синуклеин с трудом образует агрегаты.
Обработка JNPL3 тау трансгенным Было показано, что мыши с ингибитором OGA увеличивают связанный с микротрубочками белок тау O-GlcNAцилирование. Иммуногистохимический анализ ствола выявил снижение мозга образования нейрофибриллярных клубков. В анализе агрегации тиофлавина S in vitro было показано, что рекомбинантный O-GlcNAцилированный тау медленнее, чем немодифицированный тау. Аналогичные результаты получены для рекомбинантно полученной конструкции TAB1 с O-GlcNAцилированным по его немодифицированной форме.
Многие известные сайты фосфорилирования и сайты O-GlcNAцирования находятся рядом друг друга или внахлест. Белок O-GlcNAцилирование и фосфорилирование проходят как на сериновых, так и на треониновых остатках, эти посттрансляционные модификации могут регулировать друг друга. Например, в CKIIα было показано, что S347 O-GlcNAc противодействует фосфорилированию T344. Взаимное ингибирование, т.е. ингибирование фосфорилирования O-GlcNAcylation и O-GlcNAcylation фосфорилирования, наблюдалось для других белков, включая рецептор эстрогена β мыши, RNA Pol II, тау, <271.>p53, CaMKIV, p65, β-катенин и α-синуклеин. Положительная кооперативность также наблюдалась между этими двумя посттрансляционными модификациями, т.е. фосфорилирование индуцирует O-GlcNAcylation, или O-GlcNAcylation индуцирует фосфорилирование. Это было указано на MeCP2 и HDAC1. В других белках, например, кофилине, фосфорилирование и O-GlcNAцилирование, по-видимому, происходит независимо друг от друга.
В некоторых случаях изучаются терапевтические стратегии для модуляции O-GlcNAcylation с последующим эффектом фосфорилирования. Например, повышение тау-O-GlcNAclation может иметь терапевтический эффект за счет ингибирования патологического гиперфосфорирования тау-белка.
Было обнаружено, что помимо фосфорилирования, O-GlcNAc влияет на другие посттрансляционные модификации, такие как ацетилирование лизина и моноубиквитинирование.
Протеинкиназы - это ферменты, ответственные за фосфорилирование остатков серина и треонина. O-GlcNAc был идентифицирован более чем в 100 (~ 20% киназ человека кином ), и эта модификация часто занимается с изменениями активности киназы или объема субстрата киназы.
Первый отчет о киназе, непосредственно регулируемой O-GlcNAc, был опубликован в 2009 году. CaMKIV гликозилирован по множеству сайтов, хотя было обнаружено, что S189 является основным сайтом. Мутант S189A более легко активируется фосфорилированием CaMKIV T200, что позволяет предположить, что O-GlcNAc в S189 ингибирует активность CaMKIV. Гомологическое моделирование показало, что S189 O-GlcNAc может препятствовать связыванию ATP.
AMPK и OGT, как известно, модифицируют друг друга, т.е. AMPK фосфорилирует OGT, а OGT O-GlcNAцилирует AMPK. Активация AMPK рибонуклеотидом AICA связана с ядерной локализацией OGT в дифференцированных мышечных трубках скелетных мышц C2C12, что приводит к увеличению ядерной O-GlcNAc. Этот эффект не наблюдался в пролиферирующих клетках и недифференцированных миобластических клетках. Было обнаружено, что AMPK фосфорилирование OGT T444 блокирует ассоциацию OGT с хроматином и снижает H2B S112 O-GlcNAc. Было обнаружено, что сверхэкспрессия GFAT, фермента, который контролирует поток глюкозы в путь биосинтеза гексозамина, в жировой ткани мыши приводит к активации AMPK и ингибированию ACC и повышенному окислению жирных кислот. Обработка глюкозамином культивированных адипоцитов 3T3L1 показала аналогичный эффект. Точная взаимосвязь между O-GlcNAc и AMPK не была полностью установлена, поскольку в различных исследованиях сообщалось, что ингибирование OGA ингибирует активацию AMPK, ингибирование OGT также ингибирует активацию AMPK, повышающую регуляцию O-GlcNAc обработкой глюкозамином активирует AMPK, а нокдаун OGT активирует AMPK; эти результаты предполагают дополнительную непрямую связь между путями AMPK и O-GlcNAc или эффектами, специфичными для определенного типа клеток.
Было показано, что распознавание субстрата CKIIα изменяется при S347 O-GlcNAcylation.
Белковая фосфатаза 1 Субъединицы PP1β и PP1γ, как было показано, образуют функциональные комплексы с OGT. Синтетический фосфопептид мог дефосфорилироваться и O-GlcNAцилироваться иммунопреципитатом OGT. Этот комплекс называют «комплексом инь-ян», так как он заменяет фосфатную модификацию модификацией O-GlcNAc.
MYPT1 - еще одна субъединица протеинфосфатазы, которая образует комплексы с OGT и сама является O-GlcNAцилированной. MYPT1, по-видимому, играет роль в направлении OGT к определенным субстратам.
O-GlcNAцилирование белка может изменять его взаимодействие. Поскольку O-GlcNAc является высокогидрофильным, его присутствие может нарушать гидрофобные белок-белковые взаимодействия. Например, O-GlcNAc нарушает взаимодействие Sp1 с TAF II 110, а O-GlcNAc нарушает взаимодействие CREB с TAF II 130 и CRTC.
Некоторые исследования также выявили случаи, когда белок-белковые взаимодействия индуцируются O-GlcNAc. Метаболическое мечение диазирин-содержащим O-GlcNDAz применялось для идентификации белок-белковых взаимодействий, индуцированных O-GlcNAc. При использовании гликопептида-приманки, основанного примерно на консенсусной последовательности для O-GlcNAc, α-енолаза, EBP1 и 14-3-3 были идентифицированы как потенциальные O Читатели -GlcNAc. Рентгеновская кристаллография показала, что 14-3-3 распознает O-GlcNAc через амфипатическую бороздку, которая также связывает фосфорилированные лиганды. Также предполагалось, что Hsp70 действует как лектин для распознавания O-GlcNAc. Было высказано предположение, что O-GlcNAc играет роль во взаимодействии α-катенина и β-катенина.
Ко-трансляционный O- GlcNAc был идентифицирован на Sp1 и Nup62. Эта модификация подавляет ко-трансляционное убиквитинирование и, таким образом, защищает растущие полипептиды от протеасомной деградации. Наблюдались аналогичные защитные эффекты O-GlcNAc на полноразмерный Sp1. Неизвестно, универсален ли этот паттерн или применим только к конкретным белкам.
Фосфорилирование белков часто используется в качестве метки для предыдущей деградации. Белок-супрессор опухоли р53 нацелен на протеасомную деградацию через сигналосомы СОР9 -опосредованное фосфорилирование Т155. O-GlcNAцилирование p53 S149 было связано со снижением фосфорилирования T155 и защитой p53 от деградации. β-катенин O-GlcNA-цилирование конкурирует с фосфорилированием T41, которое сигнализирует β-катенину о деградации, стабилизируя белок.
O-GlcNAcylation субъединицы Rpt2 АТФазы протеасомы 26S Было показано, что ингибирует протеасомную активность. Тестирование различных пептидных последовательностей показывает, что эта модификация замедляет протеасомную деградацию гидрофобных пептидов, на деградацию гидрофильных пептидов, по-видимому, не влияет. Было показано, что эта модификация подавляет другие пути, активирующие протеасомы, такие как фосфорилирование Rpt6 с помощью цАМФ-зависимой протеинкиназы.
OGA-S локализуется в липидных каплях и имеет было предложено локально активировать протеасомы для стимулирования ремоделирования белков поверхности липидных капель.
Различные стимулы клеточного стресса были связаны с изменениями в O-GlcNAc. Обработка перекисью водорода, хлоридом кобальта (II), UVB светом, этанолом, хлоридом натрия, Тепловой шок и арсенит натрия - все это приводит к повышенному уровню O-GlcNAc. Нокаут OGT повышает чувствительность клеток к тепловому стрессу. Повышенный уровень O-GlcNAc был связан с экспрессией Hsp40 и Hsp70.
Многочисленные исследования выявили аберрантное фосфорилирование тау-белка. как признак болезни Альцгеймера. O-GlcNAцилирование бычьего тау-белка было впервые охарактеризовано в 1996 году. Последующее сообщение в 2004 году действуетало, что человеческий тау-белок также модифицируется O-GlcNAc. O-GlcNA-цилирование тау происходило, когда это происходило с образованием нейрофибриллярных клубков. Анализ мозга показал, что O-GlcNA-цилирование белка нарушено при болезни Альцгеймера, парный спиральный фрагмент-тау не распознавался традиционными методами обнаружения O-GlcNAc, что свидетельствует о патологический тау-белок нарушает O-GlcNA-цилирование по сравнению с контрольными образцов мозга. Повышение O-GlcNAцилирования тау предложено в качестве терапевтической стратегии для уменьшения фосфорилирования тау.
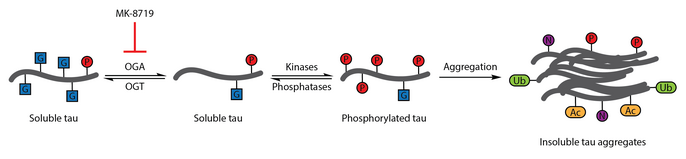 MK-8719, ингибитор OGA, подавляет агрегацию тау, повышающие уровни O-GlcNAc на тау. Посттрансляционные обозначены обозначены как G (O-GlcNAc), P (фосфорилирование), Ub (убиквитинирование), Ac (ацетилирование) и N (нитрование). Адаптировано из.
MK-8719, ингибитор OGA, подавляет агрегацию тау, повышающие уровни O-GlcNAc на тау. Посттрансляционные обозначены обозначены как G (O-GlcNAc), P (фосфорилирование), Ub (убиквитинирование), Ac (ацетилирование) и N (нитрование). Адаптировано из. Для проверки этой терапевтической гипотезы был разработан селективный и гематоэнцефалический барьер -проницаемый ингибитор OGA, тиамет-G. Обработка Thiamet-G способ увеличения тау-O-GlcNA-цитирования и подавления фосфорилирования тау-белка в культуре клеток и in vivo у здоровых крыс Sprague-Dawley. Последующее исследование показало, что обработка тиаметом-G также увеличила тау-O-GlcNAцилирование в модели трансгенных мышей JNPL3. В этой модели фосфорилирования тау-белка не подвергалось значительному влиянию тиамет-G, хотя наблюдалось уменьшение количества нейрофибриллярных клубков и более медленная потеря мотонейронов. Кроме того, было сделано, что O-GlcNAцилирование замедляет агрегацию тау in vitro.
Ингибирование OGA с помощью MK -8719 изучается в клинических испытаниях в потенциальной стратегии лечения болезни Альцгеймера и других заболеваний. таупатии, включая прогрессирующий надъядерный паралич.
Нарушение регуляции O-GlcNAc связано с пролиферацией раковых клеток и ростом опухоли.
O-GlcNAцилирование гликолитического фермента PFK1 в S529, как было обнаружено, ингибирует ферментативную активность PFK1, уменьшая гликолитический поток и перенаправляя глюкозу в сторону пентозофосфата . путь. Структурное моделирование и биохимические эксперименты показали, что O-GlcNAc на S529 ингибирует аллостерическую активацию PFK1 2,6-бисфосфатом фруктозы и олигомеризацию в активной форме. В модели на мышах мыши, вводящие клетки, экспрессирующие мутант PFK1 S529A, демонстрируют более низкий рост опухоли, чем мыши, вводящие клетки, экспрессирующие PFK1 дикого типа. Кроме того, сверхэкспрессия OGT усиливает опухоли в последней системе, но не имеет значительного влияния на систему с мутантным PFK1. Гипоксия индуцирует PFK1 S529 O-GlcNAc и увеличивает поток через пентозофосфатный путь для выработки большего количества NADPH, который поддерживает уровни глутатиона и выводит токсины на реактивные формы кислорода, улучшая рост раковых клеток. Было обнаружено, что PFK1 гликозилирован в тканях опухолей груди и легких человека. Сообщалось также, что OGT положительно регулирует HIF-1α. HIF-1α обычно разлагается в нормоксических условиях илгидроксилазами, которые используют α-кетоглутарат в качестве вспомогательного субстрата. OGT подавляет уровни α-кетоглутарата, защищая HIF-1α от протеасомной деградации pVHL и способствуя аэробному гликолизу. В отличие от предыдущего исследования PFK1, это исследование показало, что повышение уровня OGT или O-GlcNAc активировало PFK1, хотя эти два исследования согласуются с выводом о том, что уровни O-GlcNAc положительно связаны с потоком через пентозофосфатный путь. Это исследование также показало, что уменьшение O-GlcNAc избирательно убивает раковые клетки посредством ER стресса -индуцированного апоптоза.
Клеточные линии аденокарциномы поджелудочной железы (PDAC) имеют более высокий уровень O -GlcNAc по сравнению с клетками эпителия (HPDE) протока поджелудочной железы человека. Клетки PDAC имеют некоторую зависимость от O-GlcNAc для выживания, поскольку нокдаун OGT селективно ингибировал пролиферацию клеток PDAC (нокдаун OGT не оказывал значительного влияния на пролиферацию клеток HPDE), и ингибирование OGT с помощью 5S-GlcNAc показало тот же результат. Гипер-O-GlcNAцилирование в клетках PDAC оказалось антиапоптотическим, ингибируя расщепление и активацию каспазы-3 и каспазы-9. Было обнаружено, что многочисленные сайты на субъединице p65 NF-κB модифицируются O-GlcNAc динамическим образом; O-GlcNAc в p65 T305 и S319, в свою очередь, положительно регулирует другие модификации, связанные с активацией NF-κB, такие как p300 -опосредованное ацетилирование K310 и IKK -опосредованное фосфорилирование S536. Эти результаты предполагают, что NF-κB конститутивно активирует O-GlcNAc при раке поджелудочной железы.
Было обнаружено, что OGT-стабилизация EZH2 в различных клеточных линиях рака груди ингибирует экспрессию генов-супрессоров опухоли. В моделях гепатоцеллюлярной карциномы O-GlcNAc с активацией фосфорилирования HDAC1, который, в свою очередь, регулирует экспрессию регулятора клеточного цикла p21 и регулятора клеточной подвижности E-кадгерина.
Было обнаружено, что OGT стабилизирует SREBP-1 и активирует липогенез в клеточных линиях рака молочной железы. Эта стабилизация зависела от протеасомы и AMPK. Нокдаун OGT приводит к снижению ядерного SREBP-1, но протеасомное ингибирование с помощью MG132 блокировало эффект. Нокдаун OGT также усиливал взаимодействие между SREBP-1 и убиквитинлигазой E3 FBW7. AMPK активируется фосфорилированием T172 при нокдауне OGT, а AMPK фосфорилирует SREBP-1 S372, подавляя его расщепление и созревание. Нокдаун OGT оказывал влияние на уровни SREBP-1 в AMPK-нулевых клеточных линиях. В модели на мышах нокдаун OGT ингибировал рост опухоли, но сверхэкспрессия SREBP-1 частично устраняла этот эффект. Эти результаты контрастируют с результатами предыдущих исследований, в которых было обнаружено, что нокдаун / ингибирование OGT ингибирует фосфорилирование AMPK T172 и увеличивает липогенез.
В линии груди рака и простаты высокие уровни OGT и O-GlcNAc были связаны как in vitro и in vivo с процессами, связанными с прогрессированием заболеваний, например, ангиогенезом, инвазией и метастазированием. Было обнаружено, что нокдаун или ингибирование OGT подавляет фактор транскрипции FoxM1 и активирует ингибитор клеточного цикла p27 (который регулируется FoxM1-зависимой экспрессией компонента убиквитинлигазы E3 Skp2 ), вызывая остановку клеточного цикла G1. Это, по-видимому, зависело от протеасомной деградации FoxM1, поскольку экспрессия мутанта FoxM1, лишенного degron, устраняла эффекты нокдауна OGT. Было обнаружено, что FoxM1 не модифицируется напрямую с помощью O-GlcNAc, это указывает на то, что гипер-O-GlcNAцилирование регуляторов FoxM1 нарушает деградацию FoxM1. Нацеливание на OGT также снижает уровни белков, регулируемых FoxM1, связанных с инвазией и метастазированием рака (MMP-2 и MMP-9 ) и ангиогенезом (VEGF ). Также сообщалось, что модификация O-GlcNAc кофилина S108 важна для инвазии клеток рака молочной железы за счет регулирования субклеточной локализации кофилина при инвадоподиях.
Повышенное содержание O-GlcNAc имеет Был связан с диабетом.
Панкреатические β-клетки синтезируют и секретируют инсулин для регулирования уровня глюкозы в крови. Одно исследование показало, что ингибирование OGA с помощью стрептозотоцина с предыдущей обработкой глюкозамином приводит к накоплению O-GlcNAc и апоптозу в β-клетках; последующее исследование показало, что аналог стрептозотоцина на основе галактозы не может ингибировать OGA, но все же приводил к апоптозу, предполагаемая, что апоптотические эффекты стрептозотоцина не связаны напрямую с ингибированием OGA.
O-GlcNAc был предложен для ослабления сигналов инсулина. В 3T3-L1 адипоцитах ингибирование OGA с помощью PUGNAc ингибировало опосредованное инсулином поглощение глюкозы. Обработка PUGNAc также ингибировала стимулирующее инсулином фосфорилирование Akt T308 и последующее фосфорилирование GSK3β S9. В более позднем исследовании стимуляция инсулиномных клеток COS-7 вызвала локализацию OGT на плазматической мембране. Ингибирование PI3K с помощью вортманнина обращало этот эффект, предполагая зависимость от фосфатидилинозитол (3,4,5) -трифосфата. Повышение уровней O-GlcNAc путем воздействия на клетки условий с высоким уровнем глюкозы или обработки PUGNAc ингибировало стимулируемое инсулином фосфорилирование Akt T308 и активности Akt. IRS1 фосфорилирование S307 и S632 / S635, которое связано с ослабленной передачей сигналов инсулина, было усилено. Последующие эксперименты на мышах с доставкой OGT аденовирусами показали, что сверхэкспрессия OGT негативно регулирует передачу сигналов инсулина in vivo. Было обнаружено, что многие компоненты пути передачи сигналов инсулина, включая β-катенин, IR-β, IRS1, Akt, PDK1 и субъединицу p110α PI3K, быть непосредственно модифицированным O-GlcNAc. Сообщалось также, что передача сигналов инсулина приводит к фосфорилированию тирозина OGT и активации OGT, что приводит к повышению уровня O-GlcNAc.
PUGNAc также ингибирует лизосомный β-гексозаминидазы, OGA-селективный ингибитор NButGT, разработан для дальнейшего исследования между O-GlcNAc и передачей сигналов инсулина в адипоцитах 3T3 -L1. Это исследование также показало, что PUGNAc приводит к нарушению передачи сигналов инсулина, а NButGT - нет, что измеряется по изменениям в фосфорилировании Akt T308, предполагаемые, что эффекты, наблюдаемые с PUGNAc, могут быть вызваны нецелевыми эффектами, вызывающими ингибирование OGA.
Болезнь Паркинсона занимается с агрегацией α-синуклеина. Было обнаружено, что модификация α-синуклеина O-GlcNAc ингибирует его агрегацию, повышение уровня α-синуклеина O-GlcNAc исследуется как терапевтическая стратегия для лечения болезни Паркинсона.
Обработка макрофагов липополисахаридом (LPS), основным компонентом внешней мембраны грамотрицательных бактерий, приводит к повышению уровня O-GlcNAc в клетках и мышах модели. Во время инфицирования цитозольный OGT был де- S-нитрозилирован и активирован. Подавление O-GlcNAc с помощью DON ингибировало O-GlcNA-цилирование и ядерную транслокацию NF-κB, а также последующую индукцию индуцибельной синтазы оксида азота и IL-1β. Лечение ДОН также улучшало выживаемость клеток во время лечения ЛПС.
O-GlcNAc был вовлечен в вирус гриппа A (IAV) -индуцированный цитокиновый шторм. В частности, было показано, что O-GlcNA-цилирование S430 на регуляторном факторе-5 интерферона (IRF5) способствует его взаимодействию с фактором 6, действующим рецептором TNF (TRAF6), на клеточных и мышиных моделях.. TRAF6 опосредует K63-связанное убиквитинирование IRF5, которое вызывает активность IRF5 и последующего производства цитокинов. Анализ клинических образцов показал, что уровень глюкозы в крови был повышен у пациентов, инфицированных IAV, по сравнению со здоровыми людьми. У пациентов, инфицированных IAV, уровни глюкозы в крови положительно коррелировали с уровнями IL-6 и IL-8. O-GlcNAцилирование IRF5 также было относительно выше в мононуклеарных клетках периферической крови пациентов, инфицированных IAV.
Пептидные терапевтические средства, такие как привлекательные благодаря своей высокой специфичности и эффективности, но они часто имеют плохие фармакокинетические профили из-за их деградации протеазами сыворотки. Хотя O-GlcNAc обычно ассоциируется с внутриклеточными белками, было обнаружено, что сконструированные пептидные терапевтические средства, модифицированные O-GlcNAc, имеют повышенную стабильность в сыворотке на мышиной модели и имеют аналогичную активность и активность по сравнению с другими немодифицированными пептидами. Этот метод был применен для создания пептидов GLP-1 и PTH.