
Войти
| Ноцицептор | |
|---|---|
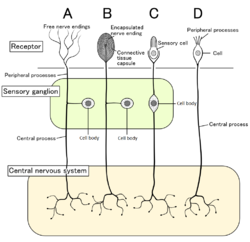 Четыре типа сенсорных нейронов и их рецепторные клетки. Ноцицепторы показаны как свободные нервные окончания тип A Четыре типа сенсорных нейронов и их рецепторные клетки. Ноцицепторы показаны как свободные нервные окончания тип A | |
| Идентификаторы | |
| MeSH | D009619 |
| Анатомическая терминология [редактирование в Викиданных ] | |
A ноцицептор («болевой рецептор») представляет собой сенсорный нейрон, который реагирует на повреждающие или потенциально повреждающие стимулы, посылая сигналы «возможной угрозы» в спинной и головной мозг. Если мозг воспринимает угрозу как достоверную, он создает ощущение боли, чтобы направить внимание на часть тела, так что можно надеяться, что угрозу можно уменьшить; этот процесс называется ноцицепцией.
Ноцицепторы были открыты Чарльзом Скоттом Шеррингтоном в 1906 году. В более ранние века ученые полагали, что животные подобны механическим устройствам, которые преобразовывают энергию сенсорных стимулов в двигательные реакции. Шеррингтон использовал множество различных экспериментов, чтобы продемонстрировать, что разные типы стимуляции афферентного нервного волокна рецептивного поля приводят к разным ответам. Некоторые интенсивные раздражители вызывают рефлекс отстранения, определенные вегетативные реакции и боль. Специфические рецепторы для этих интенсивных стимулов были названы ноцицепторами.
У млекопитающих ноцицепторы находятся в любой области тела, которая может ощущать вредные стимулы. Наружные ноцицепторы обнаруживаются в ткани, такой как кожа (кожные ноцицепторы ), роговица и слизистая оболочка. Внутренние ноцицепторы обнаруживаются во множестве органов, таких как мышцы, суставы, мочевой пузырь, висцеральные органы и пищеварительный тракт. Тела этих нейронов расположены либо в ганглиях задних корешков, либо в ганглиях тройничного нерва. Ганглии тройничного нерва - это специализированные нервы лица, тогда как ганглии задних корешков связаны с остальной частью тела. Аксоны проходят в периферическую нервную систему и заканчиваются ветвями, образуя рецептивные поля.
Ноцицепторы развиваются из стволовых клеток нервного гребня. Нервный гребень ответственен за большую часть раннего развития у позвоночных. Он особенно отвечает за развитие периферической нервной системы (ПНС). Стволовые клетки нервного гребня отделяются от нервной трубки, когда она закрывается, и ноцицепторы вырастают из дорсальной части этой ткани нервного гребня. Они формируются поздно во время нейрогенеза. Ранее формирующиеся клетки из этой области могут стать рецепторами, не воспринимающими боль, либо проприорецепторами, либо низкопороговыми механорецепторами. Все нейроны, происходящие из нервного гребня, включая эмбриональные ноцицепторы, экспрессируют TrkA, который является рецептором фактора роста нервов (NGF). Однако факторы транскрипции, которые определяют тип ноцицептора, остаются неясными.
После сенсорного нейрогенеза происходит дифференцировка и образуются два типа ноцицепторов. Они классифицируются как пептидергические или непептидергические ноцицепторы, каждый из которых выражает отдельный репертуар ионных каналов и рецепторов. Их специализация позволяет рецепторам иннервировать различные центральные и периферические мишени. Эта дифференциация происходит как в перинатальном, так и в послеродовом периоде. Непептидергические ноцицепторы выключают TrkA и начинают экспрессировать Ret, который является трансмембранным сигнальным компонентом, который обеспечивает экспрессию фактора роста, происходящего из глиальных клеток (GDNF). Этому переходу способствует Runx1, который жизненно важен для развития непептидергических ноцицепторов. Напротив, пептидергические ноцицепторы продолжают использовать TrkA, и они экспрессируют фактор роста совершенно другого типа. В настоящее время проводится множество исследований о различиях между ноцицепторами.
Периферический конец зрелого ноцицептора - это место, где вредные стимулы обнаруживаются и передаются в электрическую энергию. Когда электрическая энергия достигает порогового значения, индуцируется потенциал действия, который направляется к центральной нервной системе (ЦНС). Это приводит к череде событий, позволяющих осознать боль. Сенсорная специфичность ноцицепторов устанавливается по высокому порогу только определенных свойств раздражителей. Ноцицепторы срабатывают только тогда, когда достигается высокий порог в химической, термической или механической среде. Большинство ноцицепторов классифицируются по модальности окружающей среды, на которую они реагируют. Некоторые ноцицепторы реагируют более чем на одну из этих модальностей и, следовательно, называются полимодальными. Другие ноцицепторы не реагируют ни на одну из этих модальностей (хотя они могут отвечать на стимуляцию в условиях воспаления) и называются спящими или безмолвными.
Ноцицепторы имеют два разных типа аксонов. Первыми являются аксоны волокна Aδ. Они миелинизированы и могут позволить потенциалу действия перемещаться со скоростью около 20 метров в секунду в направлении ЦНС. Другой тип - это более медленно проводящие аксоны C волокна. Они проводят только со скоростью около 2 метров в секунду. Это происходит из-за легкого или немиелинизации аксона. В результате боль бывает двух фаз. Первая фаза обеспечивается быстропроводными волокнами Aδ, а вторая часть - волокнами (Polymodal) C. Боль, связанная с волокнами Aδ, может быть связана с первоначальной чрезвычайно острой болью. Вторая фаза - более продолжительное и немного менее интенсивное ощущение боли в результате острого повреждения. Если имеется массивный или продолжительный вход в C-волокно, происходит прогрессирующее накопление в спинном роге спинного мозга; это явление похоже на столбняк в мышцах, но называется закручиванием. Если происходит возбуждение, существует вероятность повышенной чувствительности к боли.
Тепловые ноцицепторы активируются ядовитым теплом или холодом при различных температурах. Существуют определенные преобразователи ноцицепторов, которые отвечают за то, как и если конкретное нервное окончание реагирует на тепловой стимул. Первым был обнаружен TRPV1, и его порог совпадает с температурой тепловой боли в 43 ° C. Другая температура в диапазоне от теплого до горячего определяется более чем одним каналом TRP. Каждый из этих каналов выражает определенный C-концевой домен, который соответствует чувствительности к теплу и теплу. Взаимодействие между всеми этими каналами и то, как определяется уровень температуры выше болевого порога, в настоящее время неизвестно. Холодные стимулы воспринимаются каналами TRPM8. Его С-концевой домен отличается от термочувствительных TRP. Хотя этот канал соответствует холодным стимулам, до сих пор неизвестно, способствует ли он обнаружению сильного холода. Интересное открытие, связанное с холодными стимулами, заключается в том, что тактильная чувствительность и двигательная функция ухудшаются, а восприятие боли сохраняется.
Механические ноцицепторы реагируют на избыточное давление или механическую деформацию. Они также реагируют на разрезы, повреждающие поверхность кожи. Реакция на раздражитель обрабатывается корой головного мозга как боль, как и химические и тепловые реакции. Эти механические ноцицепторы часто имеют полимодальные характеристики. Таким образом, возможно, что некоторые преобразователи тепловых раздражителей одинаковы для механических раздражителей. То же самое верно и для химических стимулов, поскольку TRPA1, по-видимому, обнаруживает как механические, так и химические изменения.
Химические ноцицепторы имеют каналы TRP, которые реагируют на широкий спектр специй. Тот, который вызывает наибольший отклик и очень широко протестирован, - это капсаицин. Другими химическими стимуляторами являются раздражители окружающей среды, такие как акролеин, химическое оружие времен Первой мировой войны и компонент сигаретного дыма. Помимо этих внешних стимуляторов, химические ноцицепторы обладают способностью обнаруживать эндогенные лиганды и определенные амины жирных кислот, которые возникают в результате изменений во внутренних тканях. Как и в тепловых ноцицепторах, TRPV1 может обнаруживать химические вещества, такие как капсаицин и токсины пауков.
Хотя каждый ноцицептор может иметь множество возможных пороговых уровней, некоторые вообще не реагируют на химические вещества., тепловые или механические раздражители, если только травма не произошла. Их обычно называют молчащими или спящими ноцицепторами, поскольку их реакция возникает только при возникновении воспаления в окружающей ткани.
Многие нейроны выполняют только одну функцию; поэтому нейроны, которые выполняют эти функции в комбинации, получают классификацию «полимодальные».
Афферентные ноцицептивные волокна (те, которые отправляют информацию в мозг, а не из него) возвращаются обратно к спинной мозг, где они образуют синапсы в его спинном роге. Это ноцицептивное волокно (расположенное на периферии) является нейроном первого порядка. клетки в спинном роге разделены на физиологически различные слои, называемые пластинками. Различные типы волокон образуют синапсы в разных слоях и используют либо глутамат, либо вещество P в качестве нейротрансмиттера. Волокна Aδ образуют синапсы в пластинках I и V, волокна C соединяются с нейронами в пластинке II, волокна Aβ соединяются с пластинками I, III и V. После достижения определенной пластинки в спинном мозге ноцицептивы первого порядка передаются нейронам второго порядка. пересекают среднюю линию передней белой комиссуры. Затем нейроны второго порядка отправляют свою информацию в таламус двумя путями: в медиально-лемнискальную систему спинного столба и переднебоковую систему. Первый предназначен больше для обычных неболевых ощущений, а боковой - для болевых ощущений. По достижении таламуса информация обрабатывается вентрально-задним ядром и отправляется в кору головного мозга в головном мозге по волокнам в задней конечности внутренней капсулы. Поскольку существует восходящий путь к мозгу, который инициирует сознательное осознание боли, существует также нисходящий путь, который модулирует болевые ощущения. Мозг может запросить выделение определенных гормонов или химических веществ, которые могут иметь обезболивающее действие, которое может уменьшить или подавить болевые ощущения. Область мозга, которая стимулирует высвобождение этих гормонов, - это гипоталамус.
. Этот эффект нисходящего торможения может быть продемонстрирован путем электрической стимуляции периакведуктальной серой области среднего мозга. Периакведуктальный серый цвет, в свою очередь, проецируется на другие области, участвующие в регуляции боли, такие как nucleus raphes magnus, который также получает аналогичные афференты от (NPG). В свою очередь, большое ядро шва проецируется в область желатинового вещества спинного рога и опосредует ощущение спиноталамических входов. Периакведуктальный серый цвет также содержит опиоидные рецепторы, что объясняет один из механизмов, с помощью которого опиоиды, такие как морфин и диацетилморфин, проявляют обезболивающее действие.
Чувствительность ноцицепторного нейрона модулируется большим количеством медиаторов во внеклеточном пространстве. Периферическая сенсибилизация представляет собой форму функциональной пластичности ноцицептора. Ноцицептор может превратиться из простого детектора вредных стимулов в детектор не вредных стимулов. В результате раздражители низкой интенсивности от регулярной активности вызывают болезненные ощущения. Это широко известно как гипералгезия. Воспаление - одна из частых причин, приводящих к сенсибилизации ноцицепторов. Обычно гипералгезия прекращается, когда воспаление уменьшается, однако иногда генетические дефекты и / или повторные травмы могут привести к аллодинии : совершенно не вредный стимул, такой как легкое прикосновение, вызывает сильную боль. Аллодиния также может быть вызвана повреждением ноцицептора в периферических нервах. Это может привести к деафферентации, то есть к развитию различных центральных процессов у выжившего афферентного нерва. В этой ситуации уцелевшие аксоны дорсальных корешков ноцицепторов могут вступать в контакт со спинным мозгом, изменяя тем самым нормальный входной сигнал.
Ноцицепция была зарегистрирована у животных, не относящихся к млекопитающим, включая рыба и широкий спектр беспозвоночных, включая пиявок, нематодных червей, морских слизней и личинок плодовых мух. Хотя эти нейроны могут иметь различные пути и отношения к центральной нервной системе, чем ноцицепторы млекопитающих, ноцицептивные нейроны у немлекопитающих часто активируются в ответ на такие же стимулы, как у млекопитающих, такие как высокая температура (40 градусов C или более), низкий pH, капсаицин., и повреждение тканей.
Из-за исторического понимания боли ноцицепторы также называют болевыми рецепторами. Хотя боль реальна, психологические факторы могут сильно влиять на субъективную интенсивность.