
Войти
| Negarnaviricota | |
|---|---|
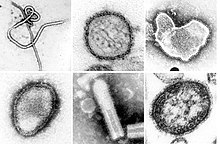 | |
| Монтаж электронных микрофотографий некоторых вирусов, относящихся к типу Negarnaviricota. Не в масштабе. Виды слева направо, сверху вниз: Эболавирус Заира, Ортохантавирус Sin Nombre, Ортопневмовирус человека, Hendra henipavirus, неустановленный рабдовирус, морбилливирус кори. | |
| Классификация вируса | |
| (без рейтинга): | Вирус |
| Область: | Рибовирия |
| Королевство: | Orthornavirae |
| Тип : | Negarnaviricota |
| Subtaxa | |
См. Текст | |
| Синонимы | |
| |
Вирусы с отрицательной РНК (-ssRNA вирусы ) представляют собой группу родственных вирусов, которые имеют одноцепочечные геномы с отрицательным смыслом, состоящие из рибонуклеиновой кислоты. У них есть геномы, которые действуют как комплементарные цепи, из которых информационная РНК (мРНК) синтезируется вирусным ферментом РНК-зависимой РНК-полимеразой (RdRp). Во время репликации вирусного генома RdRp синтезирует антигеном с положительным смыслом, который он использует в качестве матрицы для создания геномной РНК с отрицательным смыслом. Вирусы с РНК с отрицательной цепью также обладают рядом других характеристик: большинство из них содержат вирусную оболочку, которая окружает капсид, который заключает в себе вирусный геном, геномы -ssRNA вирусов обычно линейны, и это является общим для их генома. быть сегментированным.
Вирусы с отрицательной цепью РНК составляют тип Negarnaviricota в королевстве Orthornavirae и царстве рибовирии. Они произошли от общего предка, которым был вирус с двухцепочечной РНК (дцРНК), и они считаются сестринской кладой реовирусов, которые являются вирусами дцРНК. Внутри этого типа есть две основные ветви, которые образуют два подтипа: Haploviricotina, члены которого в основном несегментированы и которые кодируют RdRp, который синтезирует шапки на мРНК, и Polyploviricotina, чьи члены сегментированы и кодируют RdRp, который захватывает заглушки с мРНК хозяина. Всего распознается шесть классов этого типа.
Вирусы с РНК с отрицательной цепью тесно связаны с членистоногими и могут быть неофициально разделены на те, которые зависят от членистоногих для передачи, и те, которые произошли от вирусов членистоногих, но теперь могут воспроизводиться у позвоночных без помощи членистоногих. Известные вирусы -ssRNA, переносимые членистоногими, включают вирус лихорадки Рифт-Валли и вирус пятнистого увядания томатов. Известные вирусы -ssRNA позвоночных включают вирус Эбола, хантавирусы, вирусы гриппа, вирус лихорадки Ласса и бешенство. вирус.
Негарнавирикота берет первую часть своего названия от Латинский nega, что означает отрицательный, средняя часть РНК относится к РНК, а последняя часть, -viricota, является суффиксом, используемым для типа вируса. Подтип Haploviricotina берет первую часть своего названия, Haplo, от древнегреческого ἁπλός, что означает простой, а -viricotina - суффикс, используемый для обозначения подтипа вируса. Подтип Polyploviricotina следует той же схеме: полипло взят из древнегреческого πολύπλοκος, что означает комплекс.
 вирион вируса везикулярного стоматита (VSV) и Mononegavirales геномы
вирион вируса везикулярного стоматита (VSV) и Mononegavirales геномы Все вирусы Negarnaviricota являются вирусами с одноцепочечной РНК с отрицательным смыслом (-ssRNA). У них есть геномы, состоящие из РНК, которые являются одноцепочечными, а не двухцепочечными. Их геномы имеют отрицательный смысл, что означает, что информационная РНК (мРНК) может быть синтезирована непосредственно из генома вирусным ферментом РНК-зависимой РНК-полимеразой (RdRp), также называемой РНК-репликазой, которая кодируется всеми -ssRNA-вирусами. За исключением вирусов из рода Tenuivirus и некоторых из семейства Chuviridae, все -ssRNA вирусы имеют линейные, а не кольцевые геномы, и эти геномы могут быть сегментированными или несегментированными. Все геномы -ssРНК содержат концевые инвертированные повторы, которые представляют собой палиндромные нуклеотидные последовательности на каждом конце генома.
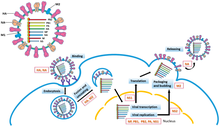 вирус гриппа цикл репликации
вирус гриппа цикл репликации Репликация геномов -ssRNA исполняется RdRp, который инициирует репликацию путем связывания с лидерной последовательностью на 3'-конце (обычно произносится как «три первичных конца») генома. Затем RdRp использует геном с отрицательным смыслом в качестве матрицы для синтеза антигенома с положительным смыслом. При репликации антигенома RdRp сначала связывается с концевой последовательностью на 3'-конце антигенома. После этого RdRp игнорирует все сигналы транскрипции на антигеноме и синтезирует копию генома, используя антигеном в качестве матрицы. Репликация выполняется, пока геном находится внутри нуклеокапсида, а RdRp раскрывает капсид и перемещается по геному во время репликации. По мере того как новые нуклеотидные последовательности синтезируются с помощью RdRp, капсидные белки собираются и инкапсидируют вновь реплицирующуюся вирусную РНК.
Транскрибирование мРНК из генома происходит по той же схеме направления, что и продуцирование антигенома. В лидерной последовательности RdRp синтезирует 5-концевую (обычно произносится как «пять простых концов») трифосфат-лидерную РНК и либо, в случае подтипа Haploviricotina, закрывает 5'-конец, либо, в случае подтипа Polyploviricotina, захватывает кэп с мРНК хозяина и прикрепляет его к вирусной мРНК, так что мРНК может транслироваться рибосомами клетки-хозяина.
после кэпинга мРНК, RdRp инициирует транскрипцию в стартовом кодоне и позже завершает транскрипцию при достижении стоп-кодона . В конце транскрипции RdRp синтезирует полиадинелированный хвост (полиА-хвост), состоящий из сотен аденинов на 3-конце мРНК, что может происходить с помощью заикания на последовательности урацилов. После конструирования хвоста полиА мРНК высвобождается с помощью RdRp. В геномах, которые кодируют более одной транскрибируемой части, RdRp может продолжить сканирование до следующего стартового кодона, чтобы продолжить транскрипцию.
Некоторые вирусы -ssRNA являются амбисенсными, что означает, что обе отрицательные геномные цепи и положительный антигеном отдельно кодируют разные белки. Чтобы транскрибировать вирусы амбисенса, выполняются два цикла транскрипции: во-первых, мРНК продуцируется непосредственно из генома; во-вторых, мРНК создается из антигенома. Все вирусы амбисенса содержат структуру шпильки для остановки транскрипции после того, как мРНК белка была транскрибирована.
 Структура перибуньявируса (слева); электронная микрофотография вируса калифорнийского энцефалита (справа)
Структура перибуньявируса (слева); электронная микрофотография вируса калифорнийского энцефалита (справа) РНК-вирусы с отрицательной цепью содержат комплекс рибонуклеопротеина, состоящий из генома и RdRp, прикрепленного к каждому сегмент генома, окруженный капсидом. Капсид состоит из белков, складчатая структура которых содержит пять альфа-спиралей в N-концевой доле (мотив 5-H) и три альфа-спирали в C-концевой доле ( 3-Н мотив). Внутри капсида геном зажат между этими двумя мотивами. За исключением семейства Aspiviridae, вирусы -ssRNA содержат внешнюю вирусную оболочку, тип липидной мембраны, окружающей капсид. Форма вирусной частицы, называемой вирионом, -ssRNA вирусов может быть нитевидной, плеоморфной, сферической или трубчатой.
Сегментация генома является важной чертой среди многих - Вирусы ssRNA и вирусы -ssRNA варьируются от геномов с одним сегментом, типичным для представителей порядка Mononegavirales, до геномов с десятью сегментами, как в случае Tilapia tilapinevirus. Нет четкой тенденции с течением времени, которая определяет количество сегментов, и сегментация генома среди -ssRNA вирусов, по-видимому, является гибким признаком, поскольку она развивалась независимо во многих случаях. Большинство представителей подтипа Haploviricotina не сегментированы, тогда как сегментация универсальна для Polyploviricotina.
 Филогенетика вирусов -ssRNA
Филогенетика вирусов -ssRNA Филогенетический анализ, основанный на RdRp, показывает, что вирусы -ssRNA происходят от общих вирусов и предком и что они, вероятно, являются сестринской кладой реовирусов, которые являются вирусами дцРНК. Внутри этого типа есть две четкие ветви, относящиеся к двум подтипам, в зависимости от того, синтезирует ли RdRp кэп на вирусной мРНК или захватывает кэп с мРНК хозяина и прикрепляет этот кэп к вирусной мРНК.
Внутри этого типа -ssRNA вирусы, которые инфицируют членистоногих, по-видимому, являются базальными и являются предками всех других -ssRNA вирусов. Членистоногие часто живут вместе большими группами, что позволяет легко передавать вирусы. Со временем это привело к тому, что -ssRNA-вирусы членистоногих приобрели высокий уровень разнообразия. Хотя членистоногие являются хозяевами большого количества вирусов, существуют разногласия по поводу степени межвидовой передачи вирусов -ssРНК членистоногих среди членистоногих.
Вирусы -ssРНК растений и позвоночных имеют тенденцию быть генетически связаны с инфицированными членистоногими вирусы. Более того, большинство -ssRNA вирусов за пределами членистоногих обнаруживается у видов, которые взаимодействуют с членистоногими. Таким образом, членистоногие служат как ключевыми хозяевами, так и векторами передачи вирусов -ssRNA. С точки зрения передачи, -ssRNA-вирусы, не относящиеся к членистоногим, можно разделить на те, которые передаются через членистоногие, и вирусы, которые могут циркулировать среди позвоночных без помощи членистоногих. Последняя группа, вероятно, произошла от первой и адаптировалась к передаче только позвоночными.
Negarnaviricota принадлежит к царству Orthornavirae, которое включает все РНК-вирусы, кодирующие RdRp, и область Riboviria, которая включает Orthornavirae, а также все вирусы, кодирующие обратную транскриптазу в королевстве Pararnavirae. Negarnaviricota содержит два подтипа, которые объединяют шесть классов, пять из которых являются монотипными вплоть до низших таксонов:
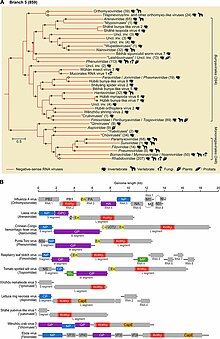 Филогенетическое дерево Negarnaviricota (вверху), геном различных членов и основные консервативные белки (внизу)
Филогенетическое дерево Negarnaviricota (вверху), геном различных членов и основные консервативные белки (внизу) Вирусы с РНК с отрицательной цепью классифицируются как Группа V в Система балтиморской классификации, которая группирует вирусы на основе их способа производства мРНК и которая часто используется вместе со стандартной систематикой вирусов, основанной на истории эволюции. Таким образом, группа V и Negarnaviricota являются синонимами.
Вирусы с отрицательной цепью РНК вызывают многие широко известные заболевания. Многие из них передаются членистоногими, включая вирус лихорадки Рифт-Валли и вирус пятнистого увядания томатов. Среди позвоночных летучие мыши и грызуны являются распространенными переносчиками многих вирусов, включая вирус Эбола и вирус бешенства, передаваемые летучими мышами и другими позвоночными животными, а также вирус лихорадки Ласса и хантавирусы, передаваемые грызунами. Вирусы гриппа распространены среди птиц и млекопитающих. Специфические для человека вирусы -ssRNA включают вирус кори и вирус эпидемического паротита.
На протяжении всей истории были известны многие заболевания, вызываемые -ssRNA-вирусами, включая хантавирусную инфекцию, корь и бешенство. В современной истории некоторые из них, такие как Эбола и грипп, вызывали вспышки смертельных заболеваний. вирус везикулярного стоматита, впервые выделенный в 1925 году и один из первых вирусов животных, подлежащих изучению, поскольку его можно было хорошо изучить в культурах клеток, был идентифицирован как вирус -ssRNA, который был уникальным в то время, потому что другие обнаруженные РНК-вирусы обладали положительным смыслом. В начале 21 века болезнь крупного рогатого скота чума крупного рогатого скота, вызванная вирусом -ssRNA чумы крупного рогатого скота, стала вторым заболеванием, которое было ликвидировано после натуральной оспы, вызванной ДНК-вирусом.
В 21 веке вирусная метагеномика стала обычным явлением для выявления вирусов в окружающей среде. Для вирусов -ssRNA это позволило идентифицировать большое количество вирусов беспозвоночных, и особенно членистоногих, что помогло получить представление об эволюционной истории вирусов -ssRNA. На основе филогенетического анализа RdRp, показывающего, что вирусы -ssRNA произошли от общего предка, Negarnaviricota и два его подтипа были установлены в 2018 году и помещены в недавно созданную область Рибовирия.
| Wikispecies содержит информацию, относящуюся к вирусу РНК с отрицательной цепью |











