
Войти
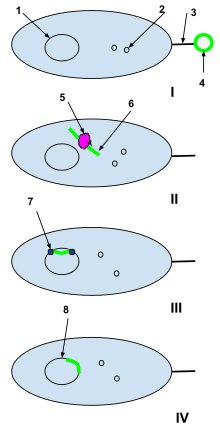 Естественная компетентность. . ДНК 1-бактериальных клеток. 2-Плазмиды бактериальных клеток. 3- Половые пили. 4-плазмида чужеродной ДНК из мертвой клетки. 5-рестрикционный фермент бактериальной клетки. 6-незакрученная чужеродная плазмида. 7-ДНК-лигаза. I:Плазмида чужеродной ДНК из мертвой клетки клетка перехватывается половыми пилями естественно компетентной бактериальной клетки.. II: Чужеродная плазмида трансдуцируется через половые пили в бактериальную клетку, где она обрабатывается ферментами рестрикции бактериальных клеток. Ферменты рестрикции разбивают чужеродную плазмиду на цепь нуклеотидов, которая может быть добавлена к бактериальной ДНК.. III: ДНК-лигаза интегрирует чужеродные нуклеотиды в ДНК бактериальной клетки.. IV: Рекомбинация завершена, и чужеродная ДНК интегрировалась в ДНК исходной бактериальной клетки и продолжит быть ее частью, когда бактериальная клетка будет реплицироваться в следующий раз.
Естественная компетентность. . ДНК 1-бактериальных клеток. 2-Плазмиды бактериальных клеток. 3- Половые пили. 4-плазмида чужеродной ДНК из мертвой клетки. 5-рестрикционный фермент бактериальной клетки. 6-незакрученная чужеродная плазмида. 7-ДНК-лигаза. I:Плазмида чужеродной ДНК из мертвой клетки клетка перехватывается половыми пилями естественно компетентной бактериальной клетки.. II: Чужеродная плазмида трансдуцируется через половые пили в бактериальную клетку, где она обрабатывается ферментами рестрикции бактериальных клеток. Ферменты рестрикции разбивают чужеродную плазмиду на цепь нуклеотидов, которая может быть добавлена к бактериальной ДНК.. III: ДНК-лигаза интегрирует чужеродные нуклеотиды в ДНК бактериальной клетки.. IV: Рекомбинация завершена, и чужеродная ДНК интегрировалась в ДНК исходной бактериальной клетки и продолжит быть ее частью, когда бактериальная клетка будет реплицироваться в следующий раз. В микробиологии, генетика, клеточная биология и молекулярная биология, компетентность - это способность клетки изменять свою генетику, принимая извлечение внеклеточной («голой») ДНК из окружающей среды в процессе, называемом трансформацией. Компетенция может быть дифференцирована между естественной компетенцией, генетически определенной способностью бактерий, которая, как считается, возникает в естественных условиях, а также в лаборатории, и индуцированной или искусственной компетенцией, которая возникает при обработке клеток в лабораторных культурах. чтобы сделать их временно проницаемыми для ДНК. Компетенция позволяет быстро адаптироваться и восстановить ДНК клетки. Эта статья в первую очередь касается естественной компетентности у бактерий, хотя также предоставляется информация об искусственной компетентности.
Природная компетентность была обнаружена Фредериком Гриффитом в 1928 году, когда он показал, что препарат убитых клеток патогенной бактерии содержит нечто, что может преобразовывать родственные непатогенные клетки в патогенные клетки. В 1944 году Освальд Эйвери, Колин МакЛауд и Маклин Маккарти продемонстрировали, что этот «трансформирующий фактор» был чистой ДНК. Это было первое убедительное доказательство того, что ДНК несет генетическую информацию клетки.
С тех пор естественная компетентность была изучена у ряда различных бактерий, в частности Bacillus subtilis, Streptococcus pneumoniae («пневмококк» Гриффита), Neisseria gonorrhoeae, Haemophilus influenzae и представители рода Acinetobacter. Области активных исследований включают механизмы транспорта ДНК, регуляцию компетентности у различных бактерий и эволюционную функцию компетентности.
В лаборатории исследователь предоставляет ДНК, часто в виде генно-инженерного фрагмента или плазмиды. Во время захвата ДНК транспортируется через клеточную мембрану (-ы) и клеточную стенку, если она присутствует. Как только ДНК оказывается внутри клетки, она может расщепляться до нуклеотидов, которые повторно используются для репликации ДНК и других метаболических функций. В качестве альтернативы он может быть рекомбинирован в геном клетки с помощью ее ферментов репарации ДНК. Если эта рекомбинация изменяет генотип клетки, говорят, что клетка трансформировалась. Искусственная компетентность и трансформация используются в качестве инструментов исследования у многих организмов (см. Трансформация (генетика) ).
Почти у всех естественно компетентных бактерий компоненты внеклеточных нитей, называемые пили IV типа (тип фимбрии ) связывают внеклеточную двухцепочечную ДНК. Затем ДНК транслоцируется через мембрану (или мембраны для грамотрицательных бактерий) через многокомпонентные белковые комплексы, управляемые разложением одной цепи ДНК.. Одноцепочечная ДНК в клетке связана с хорошо законсервированным белком, DprA, который загружает ДНК в RecA, который опосредует гомологичную рекомбинацию посредством классического пути репарации ДНК.
В лабораторных культурах естественная компетентность обычно жестко регулируется и часто запускается из-за нехватки питательных веществ или неблагоприятных условий. Однако специфические побуждающие сигналы и регуляторный механизм гораздо более разнообразны, чем механизмы усвоения, и мало что известно о регуляции компетентности в естественной среде этих бактерий. Были обнаружены факторы транскрипции, которые регулируют компетентность; примером является sxy (также известный как tfoX), который, как было обнаружено, регулируется, в свою очередь, 5'-некодирующим элементом РНК. У бактерий, способных образовывать споры, условия, вызывающие споруляцию, часто перекрываются с условиями, вызывающими компетентность. Таким образом, культуры или колонии, содержащие спорулирующие клетки, часто также содержат компетентные клетки. Недавнее исследование Süel et al. идентифицировал возбудимый основной модуль генов, который может объяснить вход в компетенцию и выход из нее, если принять во внимание клеточный шум.
Считается, что наиболее компетентные бактерии поглощают все молекулы ДНК примерно с одинаковая эффективность, но бактерии из семейств Neisseriaceae и Pasteurellaceae предпочтительно поглощают фрагменты ДНК, содержащие короткие последовательности ДНК, называемые последовательностями захвата ДНК (DUS) у Neisseriaceae и захватом сигнальная последовательность (USS) у Pasteurellaceae, которые очень часто встречаются в их собственных геномах. Геномы Neisseria содержат тысячи копий предпочтительной последовательности GCCGTCTGAA, а геномы Pasteurellacean содержат либо AAGTGCGGT, либо ACAAGCGGT.
Большинство предложений сделано для первичная эволюционная функция естественной компетентности как часть естественной бактериальной трансформации делится на три категории: (1) селективное преимущество генетического разнообразия; (2) поглощение ДНК как источника нуклеотидов (ДНК как «пища»); и (3) селективное преимущество новой цепи ДНК, способствующей гомологичной рекомбинационной репарации поврежденной ДНК (репарация ДНК). Было выдвинуто и вторичное предположение, в котором отмечалось случайное преимущество горизонтального переноса генов.
Аргументы в пользу генетического разнообразия как основной эволюционной функции пола (включая бактериальную трансформацию) являются данное Бартоном и Чарльзуортом. и Отто и Герштейн. Однако теоретические трудности, связанные с эволюцией пола, предполагают, что пол для генетического разнообразия проблематичен. В частности, что касается бактериальной трансформации, компетентность требует высокой стоимости глобального переключателя синтеза белка, например, более 16 генов, которые включаются только во время компетентности Streptococcus pneumoniae. Однако, поскольку бактерии имеют тенденцию расти в клонах, ДНК, доступная для трансформации, обычно будет иметь тот же генотип, что и клетки-реципиенты. Таким образом, всегда существует высокая стоимость экспрессии белка без увеличения разнообразия. Другие различия между компетенцией и полом были рассмотрены в моделях эволюции генов, определяющих компетенцию; Эти модели показали, что постулируемые рекомбинационные преимущества компетентности были даже более неуловимыми, чем преимущества секса.
Вторая гипотеза, ДНК как еда, основана на том факте, что клетки, которые принимают ДНК неизбежно приобретает нуклеотиды, из которых состоит ДНК, и, поскольку нуклеотиды необходимы для синтеза ДНК и РНК и дороги в синтезе, они могут вносить значительный вклад в энергетический бюджет клетки. Некоторые естественно компетентные бактерии также выделяют нуклеазы в окружающую среду, и все бактерии могут поглощать свободные нуклеотиды, которые эти нуклеазы генерируют из окружающей ДНК. Энергетика поглощения ДНК не изучена ни в одной системе, поэтому трудно сравнить эффективность секреции нуклеазы с эффективностью поглощения ДНК и внутренней деградации. В принципе, стоимость производства нуклеаз и неопределенность извлечения нуклеотидов должны быть уравновешены с энергией, необходимой для синтеза механизма поглощения и втягивания ДНК. Другими важными факторами являются вероятность того, что нуклеазы и компетентные клетки столкнутся с молекулами ДНК, относительная неэффективность поглощения нуклеотидов из окружающей среды и периплазмы (где одна цепь разрушается компетентными клетками), а также преимущества получения готовых к использованию нуклеотидных монофосфатов из другой цепи в цитоплазме. Еще одним усложняющим фактором является самоискажение систем захвата ДНК у видов из семейства Pasteurellaceae и рода Neisseria, что может отражать либо отбор по рекомбинации, либо по механически эффективному захвату.
У бактерий проблема повреждения ДНК наиболее выражена в периоды стресса, особенно окислительного стресса, который возникает в условиях скученности или голода. В таких условиях часто присутствует только одна хромосома. Открытие того, что некоторые бактерии вызывают компетентность в таких стрессовых условиях, подтверждает третью гипотезу, что существует трансформация, позволяющая восстановить ДНК. В экспериментальных тестах бактериальные клетки, подвергшиеся воздействию агентов, повреждающих их ДНК, а затем подвергшиеся трансформации, выживали лучше, чем клетки, подвергшиеся повреждению ДНК, которые не подверглись трансформации (Hoelzer and Michod, 1991). Кроме того, способность претерпевать трансформацию часто индуцируется известными повреждающими ДНК агентами (обзор Michod et al., 2008 и Bernstein et al., 2012). Таким образом, сильным краткосрочным селективным преимуществом для естественной компетентности и трансформации будет его способность способствовать гомологичной рекомбинационной репарации ДНК в условиях стресса. Такие стрессовые состояния могут возникнуть во время бактериальной инфекции восприимчивого хозяина. В соответствии с этой идеей Li et al. сообщил, что среди различных высокотрансформируемых S. pneumoniae, пригодность к назальной колонизации и вирулентность (инфекционность легких) зависят от сохранности системы компетентности.
Контраргумент был основан на отчете Редфилда 1993 года, который обнаружил, что одноцепочечные и двухцепочечные повреждения хромосомной ДНК не вызывают и не повышают компетентность или трансформацию у B. subtilis или H. influenzae, что предполагает что отбор для репарации играет незначительную роль или не играет никакой роли в эволюции компетентности у этих видов
Однако более свежие данные показывают, что способность к трансформации действительно специфически индуцируется условиями повреждения ДНК. Например, Claverys et al. в 2006 году показали, что повреждающие ДНК агенты митомицин C (агент сшивания ДНК) и фторхинолон (ингибитор топоизомеразы, вызывающий двухцепочечные разрывы) вызывают трансформацию Streptococcus pneumoniae. Кроме того, Энгельмоер и Розен в 2011 году продемонстрировали, что трансформация S. pneumoniae защищает от бактерицидного действия митомицина C. Индукция компетентности дополнительно защищает от антибиотиков каномицина и стрептомицина. Хотя ранее эти аминогликозидные антибиотики считались не повреждающими ДНК, недавние исследования, проведенные в 2012 году Foti et al. показали, что значительная часть их бактерицидной активности является результатом высвобождения гидроксильного радикала и индукции повреждений ДНК, включая двухцепочечные разрывы.
Dorer et al. В 2010 году показали, что ципрофлоксацин, который взаимодействует с ДНК-гиразой и вызывает образование двухцепочечных разрывов, индуцирует экспрессию генов компетентности в Helicobacter pylori, что приводит к усилению трансформации. В 2011 году исследования Legionella pneumophila, Charpentier et al. протестировали 64 токсичных молекулы, чтобы определить, какие из них вызывают компетентность. Только шесть из этих молекул, все агенты, повреждающие ДНК, сильно индуцировали компетентность. Этими молекулами были норфлоксацин, офлоксацин и налидиксовая кислота (ингибиторы ДНК-гиразы, которые вызывают двухцепочечные разрывы), митомицин C (который производит межцепочечные поперечные связи), бицикломицин (вызывает одно- и двухцепочечные разрывы) и гидроксимочевина (вызывает окисление оснований ДНК). Charpentier et al. также показали, что УФ-облучение индуцирует компетентность у L. pneumophila, а также предположили, что способность к трансформации развивается как ответ на повреждение ДНК.
Долгосрочное преимущество может иногда предоставляться случайными случаями горизонтального переноса гена, также называемого латеральным переносом гена (который может быть результатом негомологичная рекомбинация после индукции компетентности), которая может обеспечить устойчивость к антибиотикам или другие преимущества.
Независимо от природы отбора на компетентность, сложная природа бактериальных геномов предоставляет множество доказательств того, что горизонтальный перенос генов, вызванный компетентностью, способствует генетическому разнообразию, которое делает возможной эволюцию.