
Войти
 Ацетилхолин - природный агонист мускариновых и никотиновых рецепторов.
Ацетилхолин - природный агонист мускариновых и никотиновых рецепторов.  Мускарин - агонист, используемый для различения этих двух классов рецепторов. В теле обычно не встречается.
Мускарин - агонист, используемый для различения этих двух классов рецепторов. В теле обычно не встречается.  Атропин - антагонист.
Атропин - антагонист. Мускариновые рецепторы ацетилхолина, или mAChR, представляют собой рецепторы ацетилхолина, которые образуют рецепторные комплексы, связанные с G-белком, в клеточных мембранах определенных нейронов и других клеток. Они играют несколько ролей, в том числе действуют как главный концевой рецептор, стимулируемый ацетилхолином, высвобождаемым из постганглионарных волокон в парасимпатической нервной системе.
Мускариновые рецепторы названы так потому, что они более чувствительны к мускарину, чем к никотину. Их аналогами являются никотиновые рецепторы ацетилхолина (nAChR), ионные каналы рецепторов, которые также важны для вегетативной нервной системы. Многие лекарства и другие вещества (например, пилокарпин и скополамин ) воздействуют на эти два разных рецептора, действуя как селективные агонисты или антагонисты.
Ацетилхолин (ACh) - нейромедиатор, обнаруженный в головном мозге, нервно-мышечных соединениях и вегетативных ганглиях. Мускариновые рецепторы используются в следующих ролях:
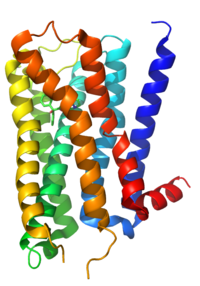 Строение мускаринового ацетилхолинового рецептора М2.
Строение мускаринового ацетилхолинового рецептора М2. ACh всегда используется как нейротрансмиттер в вегетативном ганглии. Никотиновые рецепторы постганглионарного нейрона ответственны за начальную быструю деполяризацию (Fast EPSP ) этого нейрона. Вследствие этого никотиновые рецепторы часто называют рецепторами постганглионарных нейронов ганглия. Однако последующая гиперполяризация ( IPSP ) и медленная деполяризация (Slow EPSP), которые представляют собой восстановление постганглионарного нейрона от стимуляции, на самом деле опосредуются мускариновыми рецепторами типов M 2 и M 1 соответственно (обсуждаются ниже).
Периферические вегетативные волокна (симпатические и парасимпатические волокна) анатомически классифицируются как преганглионарные или постганглионарные волокна, а затем обобщаются как адренергические волокна, выделяющие норадреналин, или холинергические волокна, оба выделяющие ацетилхолин и экспрессирующие рецепторы ацетилхолина. И преганглионарные симпатические волокна, и преганглионарные парасимпатические волокна являются холинергическими. Большинство постганглионарных симпатических волокон являются адренергическими: их нейромедиатор - норадреналин, за исключением постганглионарных симпатических волокон потовых желез, пилоэректильных мышц волос на теле и артериол скелетных мышц адреналин / норадреналин не используется.
Мозгового вещества надпочечников считается симпатической ганглий и, как и другие симпатических ганглиев, снабжена холинергических преганглионарных симпатических волокон: ацетилхолин является нейромедиатором используется в этом синапсе. В Хромаффинные клетках мозгового вещества надпочечников действуют как «модифицированные» нейроны, высвобождающего адреналин и норадреналин в кровь в качестве гормонов, а не в качестве нейротрансмиттеров. Остальные постганглионарные волокна периферической вегетативной системы относятся к парасимпатическому отделу; все являются холинергическими волокнами и используют ацетилхолин в качестве нейромедиатора.
Другая роль этих рецепторов - на стыке иннервируемых тканей и постганглионарных нейронов в парасимпатическом отделе вегетативной нервной системы. Здесь ацетилхолин снова используется в качестве нейромедиатора, а мускариновые рецепторы образуют основные рецепторы иннервируемой ткани.
Очень немногие части симпатической системы используют холинергические рецепторы. В потовых железах рецепторы мускаринового типа. Симпатическая нервная система также имеет некоторые преганглионарные нервы, заканчивающиеся хромаффинными клетками в мозговом веществе надпочечников, которые выделяют адреналин и норадреналин в кровоток. Некоторые считают, что хромаффинные клетки представляют собой модифицированные постганглионарные волокна ЦНС. В мозговом веществе надпочечников ацетилхолин используется в качестве нейромедиатора, а рецептор относится к никотиновому типу.
Соматическая нервная система использует никотиновый рецептор к ацетилхолину в нервно - мышечном соединении.
Мускариновые рецепторы ацетилхолина также присутствуют и распределены по всей местной нервной системе в постсинаптических и пресинаптических положениях. Есть также некоторые свидетельства наличия постсинаптических рецепторов на симпатических нейронах, позволяющих парасимпатической нервной системе подавлять симпатические эффекты.
Известно, что мускариновые рецепторы ацетилхолина также появляются на пресинаптической мембране соматических нейронов в нервно-мышечном соединении, где они участвуют в регуляции высвобождения ацетилхолина.
Мускариновые рецепторы ацетилхолина относятся к классу метаботропных рецепторов, которые используют G-белки в качестве сигнального механизма. В таких рецепторах сигнальная молекула ( лиганд ) связывается с рецептором, который имеет семь трансмембранных областей ; в данном случае лигандом является ACh. Этот рецептор связан с внутриклеточными белками, известными как G-белки, которые начинают информационный каскад внутри клетки.
Напротив, никотиновые рецепторы используют механизм лиганд-зависимого ионного канала для передачи сигналов. В этом случае связывание лигандов с рецептором вызывает открытие ионного канала, позволяя одному или нескольким конкретным типам ионов (например, K +, Na +, Ca 2+) диффундировать внутрь или из клетки.
Путем использования селективных радиоактивно меченных агонистов и антагонистов были определены пять подтипов мускариновых рецепторов, названных M 1 -M 5 (с использованием верхнего регистра M и нижнего индекса). Рецепторы M 1, M 3, M 5 связаны с белками Gq, а рецепторы M 2 и M 4 связаны с белками G i / o. Есть и другие системы классификации. Например, лекарство пирензепин является мускариновым антагонистом (снижает эффект ACh), который намного сильнее действует на рецепторы M 1, чем на другие подтипы. Принятие различных подтипов происходило в порядке номеров: поэтому существуют источники, которые признают только различие M 1 / M 2. Более поздние исследования склонны распознавать M 3 и самые последние M 4. [1]
Между тем генетики и молекулярные биологи охарактеризовали пять генов, которые, по-видимому, кодируют мускариновые рецепторы, названные m1-m5 (строчная буква m; без индекса). Первые четыре кода фармакологических типов M 1 -M 4. Пятый, M 5, соответствует подтипу рецептора, который до недавнего времени не обнаруживался фармакологически. Рецепторы m1 и m2 определяли на основании частичного секвенирования белков рецепторов M 1 и M 2. Остальные были найдены путем поиска гомологии с использованием биоинформатических методов.
G-белки содержат альфа-субъединицу, которая имеет решающее значение для функционирования рецепторов. Эти субъединицы могут принимать различные формы. Существует четыре широких класса форм G-белка: G s, G i, G q и G 12/13. Мускариновые рецепторы различаются по G-белку, с которым они связаны, с некоторой корреляцией в зависимости от типа рецептора. G-белки также классифицируются в соответствии с их чувствительностью к токсину холеры (CTX) и токсину коклюша (PTX, коклюш). G s и некоторые подтипы G i (G αt и G αg) восприимчивы к CTX. Только G i восприимчив к PTX, за исключением одного подтипа G i (G αz), который является невосприимчивым. Кроме того, только при связывании с агонистом те G-белки, обычно чувствительные к РТХ, также становятся восприимчивыми к СТХ.
Различные субъединицы G-белка по-разному действуют на вторичных мессенджеров, повышая регуляцию фосфолипаз, подавляя цАМФ и т. Д.
Из-за сильной корреляции с типом мускариновых рецепторов CTX и PTX являются полезными экспериментальными инструментами для исследования этих рецепторов.
| Тип | Ген | Функция | PTX | CTX | Эффекторы | Агонисты | Антагонисты |
|---|---|---|---|---|---|---|---|
| M 1 | CHRM1 |
| нет (да) | нет (да) | G q ( G i ) ( G s ): медленный EPSP. ↓ K + проводимость | ||
| M 2 | CHRM2 |
| да | нет | G i ↑ K + проводимость ↓ Ca 2+ проводимость | ||
| M 3 | CHRM3 |
| нет | нет | G q |
| |
| M 4 | CHRM4 | да | ? | G i ↑ K + проводимость ↓ Ca 2+ проводимость | |||
| M 5 | CHRM5 |
| нет | ? | G q |
Этот рецептор опосредует медленный ВПСП в ганглии постганглионарного нерва, часто встречается в экзокринных железах и в ЦНС.
Он преимущественно связан с G-белками класса G q, которые используют активацию фосфолипазы C и, следовательно, трифосфата инозита и внутриклеточного кальция в качестве сигнального пути. Связанный таким образом рецептор не будет восприимчив к CTX или PTX. Однако также было показано, что G i (вызывающий снижение цАМФ ниже по течению) и G s (вызывающий увеличение цАМФ) также участвуют во взаимодействиях в определенных тканях и, следовательно, будут восприимчивы к PTX и CTX, соответственно.
Мускариновые рецепторы M 2 расположены в сердце и легких. В сердце они замедляют частоту сердечных сокращений ниже нормального исходного синусового ритма, замедляя скорость деполяризации. У людей в условиях покоя активность блуждающего нерва преобладает над симпатической. Следовательно, ингибирование рецепторов m2 (например, атропином) вызовет учащение пульса. Они также умеренно снижают сократительные силы предсердной сердечной мышцы и снижают скорость проводимости атриовентрикулярного узла (АВ-узла). Он также служит для небольшого уменьшения сократительной силы желудочковой мышцы.
Мускариновые рецепторы M 2 действуют через рецептор типа G i, который вызывает снижение цАМФ в клетке, ингибирование потенциал-управляемых каналов Ca 2+ и увеличение оттока K +, что в целом приводит к эффектам ингибиторного типа.
Мускариновые рецепторы M 3 расположены во многих частях тела. Они расположены в гладких мышцах кровеносных сосудов, а также в легких. Поскольку рецептор M 3 связан с G q и опосредует увеличение внутриклеточного кальция, он обычно вызывает сокращение гладкой мускулатуры, такое как наблюдаемое во время сужения бронхов и опорожнения мочевого пузыря. Однако, что касается сосудистой сети, активация M 3 на эндотелиальных клетках сосудов вызывает повышенный синтез оксида азота, который диффундирует к соседним клеткам гладких мышц сосудов и вызывает их расслабление, тем самым объясняя парадоксальный эффект парасимпатомиметиков на тонус сосудов и бронхиолярный тонус. Действительно, прямая стимуляция гладкой мускулатуры сосудов, M 3 опосредует сужение сосудов при заболеваниях, при которых нарушен эндотелий сосудов. Рецепторы M 3 также расположены во многих железах, которые помогают стимулировать секрецию, например, в слюнных железах, а также в других железах тела.
Подобно мускариновому рецептору M 1, рецепторы M 3 представляют собой G-белки класса G q, которые активируют фосфолипазу C и, следовательно, трифосфат инозита и внутриклеточный кальций в качестве сигнального пути.
Рецепторы M 4 находятся в ЦНС.
Рецепторы M 4 работают через рецепторы G i, уменьшая цАМФ в клетке и, таким образом, оказывают в целом ингибирующие эффекты. Возможный бронхоспазм может возникнуть при стимуляции мускариновыми агонистами.
Расположение рецепторов M 5 не очень хорошо известно.
Подобно мускариновым рецепторам M 1 и M 3, рецепторы M 5 связаны с G-белками класса G q, которые активируют фосфолипазу C и, следовательно, инозитолтрифосфат и внутриклеточный кальций в качестве сигнального пути.
Лиганды, нацеленные на mAChR, которые в настоящее время одобрены для клинического использования, включают неселективные антагонисты для лечения болезни Паркинсона, атропин (для расширения зрачка ), скополамин (используется для предотвращения укачивания ) и ипратропий (используется для лечения ХОБЛ )..