
Войти
| Моноцеркомоноиды | |
|---|---|
 | |
| Monocercomonoides melolanthae | |
| Научная классификация | |
| Домен: | Эукариоты |
| (без рейтинга): | Экскавата |
| Тип: | Метамонада |
| Класс: | Анаэромонада |
| Порядок: | Оксимонадида |
| Семья: | Полимастигиды |
| Род: | Моноцеркомоноиды Трэвис, 1932 год. |
| Разновидность | |
| |
Monocercomonoides является родом из жгутиковых Excavata, принадлежащий к порядку Oxymonadida. Он был установлен Бернардом В. Трэвисом и впервые описан как «жгутики полимастигинид, имеющие три передних жгутика и один задний, берущий начало в единственной базальной грануле, расположенной перед расположенным спереди ядром, и более или менее четко выраженный аксостиль. ". Это первый род эукариот, в котором полностью отсутствуют митохондрии и все основные белки, отвечающие за функцию митохондрий. В роду также отсутствуют какие-либо другие органеллы, связанные с митохондриями (MRO), такие как гидрогеносомы или митосомы. Данные свидетельствуют о том, что отсутствие митохондрий не является наследственным признаком, а скорее связано с вторичной потерей. Monocercomonoides sp. Было обнаружено, что для получения энергии за счет ферментативного действия питательных веществ, поглощаемых из окружающей среды. Род заменилпуть сборки кластера железо-сера цитозольной системой мобилизации серы, вероятно, полученной путем горизонтального переноса генов от эубактерий общего предка оксимонад. Эти организмы важны, потому что они опровергают представление о том, что у эукариот должны быть митохондрии для правильного функционирования. Генома из Monocercomonoides имеет приблизительно 75 миллионов пар оснований (75 МВР), с 16629 предсказанных белок-кодирующих генов.
Виды Monocercomonoides - это облигатные симбионты животных, обитающие в пищеварительном тракте насекомых, амфибий, рептилий и млекопитающих. Monocercomonoides обычны в отрядах прямокрылых и жесткокрылых насекомых. Вид Monocercomonoides qadrii обнаружен в прямой кишке личинки навозного жука ( Oryctes rhinoceros ). Следующие виды M. caviae, M. wenrichi, M. quadrifunilis и M. exilis обнаружены в слепой кишке морских свинок, а M. caprae обнаружены в рубце коз. В исследовании Karnkowska (2016) Monocercomonoides были обнаружены в кишечнике шиншиллы, принадлежавшей одному из сотрудников лаборатории. Исследователи обнаружили организмы в кишечнике хозяев шиншиллы, где много питательных веществ, но мало кислорода. Организм использует ферменты, обнаруженные в его цитоплазме, для расщепления пищи и выработки энергии, поскольку митохондрии и кислород отсутствуют. Было отмечено, что Monocercomonoides заглатывают бактерии или древесину и питаются пиноцитозом, однако по стилю кормления были проведены ограниченные исследования.
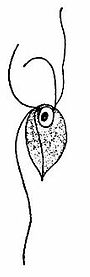
Моноцеркомоноиды - это небольшие свободно плавающие одноклеточные организмы размером от 5 до 12 мкм в длину и от 4,5 до 14,5 мкм в ширину. Тело может быть яйцевидным, грушевидным, сферическим или субсферическим; однако у них нет опор и небольшой аксостиль. Axostyle представляет собой единый, стягивает придаток из микротрубочек, который берет свое начало от заднего конца preaxostyle, и расположено вблизи задней пару базальных тел (известные как базальное тельце в пожилом цитологической литературы). Цитоскелет состоит из четырех базальных тел, передней пары и задней пары. Преаксостиль проходит между двумя парами базальных тел и состоит из широкого изогнутого листа микротрубочек. Внутренняя поверхность листа микротрубочек прилегает к паракристаллическому волокну (толщиной около 50 нм), что является общей характеристикой всех оксимонад. Monocercomonoides sp. имеет четыре жгутика, которые образуют две пары и отходят от каждого базального тела на переднем конце. Три из четырех жгутиков примерно одинаковой длины (9,5-18 мкм), а четвертый задний жгутик немного длиннее, его размер составляет 9,0-24,5 мкм. Жгутики имеют биение и используются для быстрого движения. Проксимальная часть длинного жгутика может прилипать к пленке, что заставляет ее тянуться кзади. Задний жгутик всегда направлен назад и прикреплен к телу на значительном расстоянии (6-9 мкм) дополнительной нитью, называемой funis. Есть от одной до четырех нитей (ребристых стриктур), отходящих назад под поверхность тела. У некоторых паразитов жгутики оканчиваются акронемами. Ядро обычно расположено около переднего конца тела и содержит центральную эндосому, окруженную гранулами хроматина, у некоторых видов есть пелта-подобные структуры под ядром. Цитоплазма гранулированная, с вакуолями или без них. Электронно-микроскопическое изображение Monocercomonoides показало, что внутриклеточная морфология лишена какого-либо аппарата Гольджи, митохондрий или потенциальных гомологов этих двух; Белки, ассоциированные с Гольджи, обнаружены, а митохондриальные - нет.
Monocercomonoides sp. это первый эукариот, у которого обнаружено отсутствие каких-либо следов митохондрий. У всех других эукариот, у которых, по-видимому, отсутствуют митохондрии, есть ядерная ДНК, которая содержит некоторые из генов, необходимых для сборки митохондрий, но такие гены отсутствуют у Monocercomonoides. В нем также отсутствуют какие-либо гены, обычно встречающиеся в митохондриальной ДНК, и гены, используемые для выработки энзимов, выделяющих энергию, присутствующих в митохондриях. Моноцеркомоноиды способны получать некоторую энергию из глюкозы с помощью анаэробных метаболических путей, которые действуют в цитоплазме, однако большая часть его энергии получается с помощью ферментов, расщепляющих аминокислоту аргинин.
Каждая молекула глюкозы, катаболизированная в Monocercomonoides, дает меньше АТФ по сравнению с митохондриальными эукариотами, которые используют цикл трикарбоновых кислот и цепь переноса электронов. Чтобы помочь в сохранении энергии, Monocercomonoides адаптировал альтернативные гликолитические ферменты. Четыре альтернативных гликолитических фермента включают пирофосфат-фруктозо-6-фосфатфосфотрансферазу (PFP), фруктозобисфосфатальдолазу класса II (FBA, класс II), 2,3-бисфосфоглицерат-независимую фосфоглицератмутазу (iPGM) и пируват-фосфатдикиназу. Глюкозо-6-фосфат-изомераза (GPI), по прогнозам, входит в состав Monocercomonoides, поскольку она повсеместно распространена среди эукариот, бактерий и некоторых архей и играет важную роль в катаболическом гликолизе, но еще не обнаружена. Большинство гликолитических ферментов являются стандартными эукариотическими версиями, что делает метаболический путь моноцеркомоноидов мозаикой, аналогичной метаболическим путям других анаэробов.
Добавление PPDK к превращению фосфоенолпирувата в пируват (обычно катализируемое исключительно пируваткиназой) оказывает сильное влияние на консервацию АТФ. И PFP, и PPDK полагаются на неорганический фосфат (PPi) в качестве донора фосфата; поэтому вместо гидролиза АТФ выход АТФ увеличивается за счет использования побочного продукта анаболических процессов клетки в качестве источника энергии. Эти реакции могут способствовать большей консервации АТФ и регулированию гликолиза из-за обратимой природы PPDK и использования неорганического фосфата, где пируваткиназа катализирует только прямую реакцию.
В дополнение к регулируемому гликолизу моноцеркомоноиды содержат ферменты, необходимые для пути аргининдезиминазы (деградации). Путь аргининдезиминазы можно использовать для продукции АТФ, как в Giardia Кишечник и Trichomonas vaginalis. У G. кишечника (анаэробный одноклеточный эукариот) этот путь продуцирует в восемь раз больше АТФ, чем метаболизм сахара, и аналогичный выход ожидается у моноцеркомоноидов, но еще не подтвержден.
Кластеры железо-сера - важные белковые компоненты, которые синтезируются митохондриями. Основная функция этих небольших неорганических простетических групп - опосредование транспорта электронов, что делает их ключевой частью фотосинтеза, дыхания, репликации / репарации ДНК и регуляции экспрессии генов. В эукариотических клетках обычным путем синтеза кластера Fe-S является ISC (железо-серный кластер). В цитозоле цитозольная сборка железо-серных кластеров (CIA) формирует Fe-S-кластерные белки, которые ответственны за созревание ядерных белков Fe-S. CIA уникален для эукариот и не имеет прокариотических гомологов. Считается, что митохондриальный путь ISC необходим для функционирования CIA, поскольку он синтезирует и транспортирует нехарактеризованный серосодержащий предшественник в цитозоль и является основной причиной удержания митохондриальных органелл у анаэробных эукариот. Род Monocercomonoides содержит пути CIA, но полностью лишен пути ISC вместе с любыми митохондриальными белками. Род содержит сокращенную версию пути SUF (фактора утилизации серы), имея только три белка - SufB, SufC и SufU. Путь SUF - это известный путь прокариот, и считается, что гены, используемые для построения системы SUF у Monocercomonoides, должны были происходить от прокариот. Однако было обнаружено, что белки SUF Monocercomonoides не связаны с гомологами пластид или любыми другими микробными эукариотами. Было высказано предположение, что этот путь был получен от эубактерий путем горизонтального переноса генов (HGT) у общего предка Monocercomonoides и Paratrismastrix (родственный таксон оксимонад). Генетическое приобретение не было продемонстрировано, несмотря на предположение, что оно должно было произойти.
Monocercomonoides не содержат заметных признаков того, что митохондрии / электростанция клетки когда-либо были частью организма. Однако, поскольку широко распространено мнение, что все эукариоты имеют общего предка, который развил митохондрии, считается, что митохондрии должны были когда-то присутствовать у предков оксимонад, а затем вторично утрачены. Род амитхондрий демонстрирует, что митохондрии не являются абсолютно необходимыми для жизни эукариотической клетки.
Отсутствие митохондрий или каких-либо связанных с митохондриями органелл у Monocercomonoides подтверждается последовательностью его генома. Полный анализ последовательности генома Monocercomonoides sp. PA203 из Chinchilla lanigera. Предполагаемый размер генома составляет ~ 75 МБ, а количество предсказанных генов, кодирующих белок, составляет 16629. Поиск по гомологии показывает отсутствие генов, которые кодируют механизмы импорта митохондрий, белки транспорта метаболитов и кластеры железо-сера. Кроме того, отсутствие целевых важных генов и генов, кодирующих белки митохондриальной мембраны, было выявлено при поиске специфических N-концевых и C-концевых последовательностей. Гены, которые обычно кодируются в митохондриальных геномах (мтДНК), не были обнаружены среди собранных каркасов, что позволяет предположить, что у Monocercomonoides отсутствует мтДНК. Гены 18S РНК были секвенированы, и было обнаружено, что их длина составляет 2927 нуклеотидов, и они являются одними из самых длинных из известных. Некоторые расширения были специфичны для Monocercomonoides, но многие были похожи на расширения других родов оксимонад, но значительно длиннее. Сравнения генов, кодирующих альфа-тубулина, бета-тубулина, γ-тубулина, EF-1, EF-2, cytHSP70, убиквитин, 18S рРНК, и HSP90 позволяют размещать oxymonads вблизи дипломонады и трихомонады, с Monocercomonoides и Streblomastix составление ветвь оксимонад.