
Войти
 Микрофотография, на которой показаны конденсированные хромосомы в синем, кинетохорах в розовом и микротрубочках в зеленом во время метафазы митоза
Микрофотография, на которой показаны конденсированные хромосомы в синем, кинетохорах в розовом и микротрубочках в зеленом во время метафазы митоза В клеточной биологии, веретенообразный аппарат (или митотическое веретено ) относится к цитоскелетной структуре эукариотических клеток, которая формируется во время клетки деление для разделения сестринских хроматид между дочерними клетками. Он называется митотическим веретеном во время митоза, процессом, который производит генетически идентичные дочерние клетки, или мейотическим веретеном во время мейоза, процессом, который производит гаметы с половиной числа хромосом родительской клетки.
Помимо хромосом, веретенообразный аппарат состоит из сотен белков. Микротрубочки составляют самые многочисленные компоненты механизма.
 На этой диаграмме изображена организация типичного митотического веретена, обнаруженного в клетках животных. Хромосомы прикрепляются к кинетохоре микротрубочкам через мультипротеиновый комплекс, называемый кинетохорой. Полярные микротрубочки переплетаются в средней зоне веретена и раздвигают полюса веретена через моторные белки. Астральные микротрубочки прикрепляют полюса веретена к клеточной мембране. Полимеризация микротрубочек зарождается в центре организации микротрубочек.
На этой диаграмме изображена организация типичного митотического веретена, обнаруженного в клетках животных. Хромосомы прикрепляются к кинетохоре микротрубочкам через мультипротеиновый комплекс, называемый кинетохорой. Полярные микротрубочки переплетаются в средней зоне веретена и раздвигают полюса веретена через моторные белки. Астральные микротрубочки прикрепляют полюса веретена к клеточной мембране. Полимеризация микротрубочек зарождается в центре организации микротрубочек.Присоединение микротрубочек к хромосомам опосредуется кинетохорами, которые активно контролируют формирование веретена и предотвращают преждевременную анафазу начало. Полимеризация микротрубочек и деполимеризация динамически приводят к конгрессу хромосом. Деполимеризация микротрубочек вызывает напряжение на кинетохорах; биполярное прикрепление сестринских кинетохор к микротрубочкам, исходящим от противоположных полюсов клетки, объединяет противодействующие силы натяжения, выравнивая хромосомы на экваторе клетки и настраивая их для сегрегации дочерними клетками. Как только каждая хромосома становится биориентированной, начинается анафаза и cohesin, который объединяет сестринские хроматиды, разрывается, что позволяет переходить сестринские хроматиды к противоположным полюсам.
Аппарат клеточного веретена включает веретено микротрубочки, ассоциированные белки, которые включают молекулярные моторы кинезин и динеин, конденсированные хромосомы и любые центросомы или звездочки, которые могут присутствовать на полюсах веретена в зависимости от типа клетки. Шпиндельный аппарат имеет в поперечном сечении нечетко эллипсоид и сужается на концах. В широкой средней части, известной как средняя зона веретена, антипараллельные микротрубочки связаны в пучки с помощью кинезинов. На заостренных концах, известных как полюса веретена, микротрубочки зарождаются с помощью центросом в большинстве клеток животных. Ацентросомные или анастральные веретена лишены центросом или звездочек на полюсах веретена, соответственно, и встречаются, например, во время мейоза самок у большинства животных. В этом случае градиент Ran GTP является основным регулятором организации и сборки микротрубочек веретена. У грибов веретена образуются между телами полюсов веретена, встроенными в ядерную оболочку, которая не разрушается во время митоза.
Динамическое удлинение и укорочение микротрубочек веретена посредством процесса, известного как динамическая нестабильность, в значительной степени определяет форму митотическое веретено и способствует правильному выравниванию хромосом в средней зоне веретена. Связанные с микротрубочками белки (MAP) связываются с микротрубочками в средней зоне и полюсах веретена, чтобы регулировать их динамику. γ-тубулин - это специализированный вариант тубулина, который собирается в кольцевой комплекс, называемый γ-TuRC, который зарождается полимеризацией α / β тубулина гетеродимеров в микротрубочки. Привлечение γ-TuRC в перицентросомальную область стабилизирует минус-концы микротрубочек и закрепляет их около центра организации микротрубочек. Связанный с микротрубочками белок Augmin действует совместно с γ-TURC, чтобы зародить новые микротрубочки из существующих микротрубочек.
Растущие концы микротрубочек защищены от катастрофы действием белков отслеживания микротрубочек на плюс-конце (+ TIPs), чтобы способствовать их ассоциации с кинетохорами в средней зоне. CLIP170, как было показано, локализуется вблизи плюс-концов микротрубочек в клетках HeLa и накапливается в кинетохорах во время прометафазы. Хотя как CLIP170 распознает плюс-концы, остается неясным, было показано, что его гомологи защищают от катастроф и способствуют спасению, подтверждая роль CLIP170 в стабилизации плюс-концов и, возможно, опосредуя их прямое прикрепление к кинетохорам. CLIP-ассоциированные белки, такие как CLASP1 у людей, также, как было показано, локализуются на плюс-концах и на внешней кинетохоре, а также модулируют динамику микротрубочек кинетохор (Maiato 2003). Гомологи CLASP у Drosophila, Xenopus и дрожжей необходимы для правильной сборки веретена; у млекопитающих CLASP1 и CLASP2 вносят вклад в правильную сборку веретена и динамику микротрубочек в анафазе. Полимеризация плюсовых концов может дополнительно замедляться белком EB1, который напрямую связывает растущие концы микротрубочек и координирует связывание других + TIP.
Действие этих стабилизирующих микротрубочки белков противодействует ряду микротрубочек. -деполимеризующие факторы, которые позволяют динамическое ремоделирование митотического веретена, способствуя конгрессии хромосом и достижению биполярности. Суперсемейство MAP кинезин -13 содержит класс моторных белков, направленных на положительный конец, с ассоциированной активностью деполимеризации микротрубочек, включая хорошо изученные MCAK млекопитающих и Xenopus XKCM1. MCAK локализуется на растущих кончиках микротрубочек на кинетохорах, где он может вызвать катастрофу в прямой конкуренции со стабилизирующей активностью + TIP. Эти белки используют энергию гидролиза АТФ, чтобы вызвать дестабилизирующие конформационные изменения в структуре протофиламента, которые вызывают высвобождение кинезина и деполимеризацию микротрубочек. Потеря их активности приводит к многочисленным митотическим дефектам. Дополнительные дестабилизирующие микротрубочки белки включают Op18 / stathmin и катанин, которые играют роль в ремоделировании митотического веретена, а также в содействии сегрегации хромосом во время анафазы.
Активность этих MAPs. тщательно регулируются для поддержания надлежащей динамики микротрубочек во время сборки веретена, причем многие из этих белков служат субстратами Aurora и Polo-like kinase.
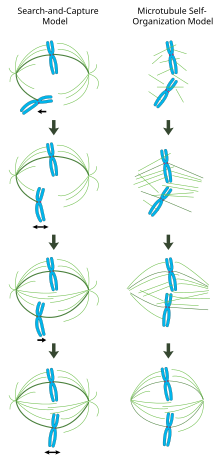 В модели «поиска и захвата», опосредованной центросомами (слева), микротрубочки, образовавшиеся из центросом, случайно контактируют с хромосомами и стабилизируются на кинетохорах с образованием веретена. В модели «самоорганизации», опосредованной хроматином (справа), микротрубочки зарождаются вокруг митотического хроматина и организуются в биполярный массив моторными белками.
В модели «поиска и захвата», опосредованной центросомами (слева), микротрубочки, образовавшиеся из центросом, случайно контактируют с хромосомами и стабилизируются на кинетохорах с образованием веретена. В модели «самоорганизации», опосредованной хроматином (справа), микротрубочки зарождаются вокруг митотического хроматина и организуются в биполярный массив моторными белками. В правильно сформированном митотическом веретене би-ориентированные хромосомы выровнены. вдоль экватора клетки с микротрубочками веретена, ориентированными примерно перпендикулярно хромосомам, их плюс-концы заключены в кинетохоры, а их минус-концы закреплены на полюсах клетки. Точная ориентация этого комплекса требуется для обеспечения точной сегрегации хромосом и определения плоскости деления клетки. Однако остается неясным, как веретено становится организованным. В этой области преобладают две модели, которые являются синергетическими и не исключающими друг друга. В модели поиска и захвата веретено преимущественно организовано за счет разделения полюсов центросомных центров организации микротрубочек (MTOCs). Микротрубочки веретена исходят из центросом и «ищут» кинетохоры; когда они связываются с кинетохорой, они стабилизируются и оказывают давление на хромосомы. В альтернативной модели самосборки микротрубочки подвергаются ацентросомному зарождению среди конденсированных хромосом. Ограниченные размерами клетки, латеральными ассоциациями с антипараллельными микротрубочками через моторные белки и концевыми присоединениями к кинетохорам, микротрубочки естественным образом принимают веретенообразную структуру с хромосомами, выровненными вдоль экватора клетки.
В этой модели микротрубочки зарождаются в центрах организации микротрубочек и подвергаются быстрому росту и катастрофе, чтобы «искать» в цитоплазме кинетохоры. Как только они связываются с кинетохорами, они стабилизируются, а их динамика снижается. Недавно моноориентированная хромосома колеблется в пространстве около полюса, к которому она прикреплена, пока микротрубочка с противоположного полюса не свяжет сестринскую кинетохору. Это второе прикрепление дополнительно стабилизирует прикрепление кинетохор к митотическому веретену. Постепенно двунаправленная хромосома тянется к центру клетки до тех пор, пока натяжение микротрубочек не уравновесится с обеих сторон центромеры ; затем сжатая хромосома колеблется на метафазной пластинке, пока начало анафазы не высвободит сцепление сестринских хроматид.
В этой модели центры организации микротрубочек локализованы на полюсах клетки, их разделение обусловлено полимеризацией микротрубочек и «скольжением» антипараллельных микротрубочек веретена относительно друг друга в средней зоне веретена, опосредованной биполярным положительным концом. -направленные кинезины. Такие силы скольжения могут объяснять не только разделение полюсов веретена на ранних этапах митоза, но также удлинение веретена во время поздней анафазы.
В отличие от механизма поиска и захвата, в котором центросомы в значительной степени диктуют организацию митотического веретена, эта модель предполагает, что микротрубочки являются зарождаются ацентросомно около хромосом и спонтанно собираются в антипараллельные пучки и принимают веретенообразную структуру. Классические эксперименты Хилда и Карсенти показывают, что функциональные митотические веретена и ядра образуются вокруг покрытых ДНК шариков, инкубированных в экстрактах яиц Xenopus, и что биполярные массивы микротрубочек образуются в отсутствие центросом и кинетохор. В самом деле, также было показано, что лазерная абляция центросом в клетках позвоночных не ингибирует ни сборку веретена, ни сегрегацию хромосом. Согласно этой схеме форма и размер митотического веретена являются функцией биофизических свойств перекрестно-связывающих моторных белков.
Фактор обмена гуаниновых нуклеотидов для малой GTPase Ran (регулятор конденсации хромосом 1 или RCC1 ) прикрепляется к нуклеосомам через коровые гистоны H2A и H2B. Таким образом, вокруг митотического хроматина образуется градиент GTP-связанного Ran. Стеклянные шарики, покрытые RCC1, вызывают зарождение микротрубочек и формирование биполярного веретена в экстрактах яиц Xenopus, показывая, что одного градиента Ran GTP достаточно для сборки веретена. Градиент запускает высвобождение факторов сборки веретена (SAFs) от ингибирующих взаимодействий через транспортные белки importin β / α. Несвязанные SAF затем способствуют зарождению микротрубочек и стабилизации вокруг митотического хроматина, а биполярность веретена организована моторными белками микротрубочек.
Сборка веретена в значительной степени регулируется событиями фосфорилирования, катализируемыми митотическими процессами. киназы. Циклинзависимые киназные комплексы (CDK) активируются митотическими циклинами, трансляция которых увеличивается во время митоза. CDK1 (также называемая CDC2) считается основной митотической киназой в клетках млекопитающих и активируется циклином B1. Киназы Aurora необходимы для правильной сборки и разделения веретена. Aurora A ассоциируется с центросомами и, как полагают, регулирует митотический вход. Аврора B является членом хромосомного комплекса-пассажира и опосредует прикрепление хромосомы к микротрубочке и сцепление сестринских хроматид. Поло-подобная киназа, также известная как PLK, особенно PLK1, играет важную роль в поддержании веретена, регулируя динамику микротрубочек.
К концу ДНК При репликации сестринские хроматиды связаны вместе в аморфную массу запутанной ДНК и белка, которую было бы практически невозможно разделить на каждую дочернюю клетку. Чтобы избежать этой проблемы, митотическое вступление запускает драматическую реорганизацию дублированного генома. Сестринские хроматиды распутаны и отделены друг от друга. Хромосомы также укорачиваются в длину, до 10 000 раз в клетках животных, в процессе, называемом конденсацией. Конденсация начинается в профазе, и хромосомы максимально уплотняются в стержневидные структуры к тому времени, когда они выравниваются в середине веретена в метафазе. Это придает митотическим хромосомам классическую Х-образную форму, наблюдаемую в кариотипах, где каждая конденсированная сестринская хроматида связана по своей длине белками когезин и соединена, часто около центра, на центромера.
Хотя эти динамические перестройки жизненно важны для обеспечения точной и высокоточной сегрегации генома, наше понимание структуры митотической хромосомы остается в значительной степени неполным. Однако были идентифицированы несколько специфических молекулярных игроков: Топоизомераза II использует гидролиз АТФ, чтобы катализировать распутывание цепочек ДНК, способствуя разрешению сестринских хроматид. Конденсины представляют собой комплексы из 5 субъединиц, которые также используют АТФ-гидролиз для ускорения конденсации хромосом. Эксперименты с экстрактами яиц Xenopus также показали, что линкер гистон H1 является важным регулятором уплотнения митотических хромосом.
Завершение формирования веретена имеет решающее значение Точка перехода в клеточном цикле называется контрольной точкой сборки шпинделя. Если к моменту этой контрольной точки хромосомы не прикреплены должным образом к митотическому веретену, наступление анафазы будет отложено. Отказ этой контрольной точки сборки веретена может привести к анеуплоидии и может быть вовлечен в старение и формирование рака.
 Рисунок делящейся клетки эпителия, окруженной тканью эпителия. Шпиндельный аппарат вращается внутри ячейки. Вращение является результатом притяжения астральных микротрубочек к трехклеточным соединениям (TCJ), сигнальным центрам, локализованным в областях, где встречаются три клетки.
Рисунок делящейся клетки эпителия, окруженной тканью эпителия. Шпиндельный аппарат вращается внутри ячейки. Вращение является результатом притяжения астральных микротрубочек к трехклеточным соединениям (TCJ), сигнальным центрам, локализованным в областях, где встречаются три клетки. Ориентация деления клеток имеет большое значение для архитектуры ткани, судьбы клеток и морфогенез. Клетки имеют тенденцию делиться вдоль своей длинной оси в соответствии с так называемым правилом Гертвига. Ось деления клеток определяется ориентацией веретенообразного аппарата. Клетки делятся по линии, соединяющей две центросомы веретенообразного аппарата. После образования веретенообразный аппарат совершает вращение внутри ячейки. Астральные микротрубочки, происходящие из центросом, достигают клеточной мембраны, где они тянутся к определенным кортикальным подсказкам. In vitro распределение кортикальных ключей устанавливается с помощью клейкого рисунка. Сигналы полярности in vivo определяются локализацией локализованных в вершинах клеток. Пространственное распределение корковых ключей приводит к силовому полю, которое определяет окончательную ориентацию веретенообразного аппарата и последующую ориентацию деления клеток.