
Войти
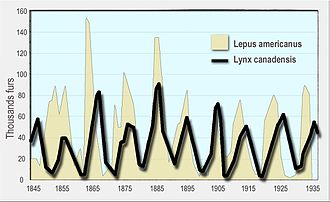 Количество меха зайца-снегоступа (желтый, на заднем плане) и канадской рыси (черная линия, передний план), проданных компании Гудзонова залива. Канадские рыси едят зайцев на снегоступах.
Количество меха зайца-снегоступа (желтый, на заднем плане) и канадской рыси (черная линия, передний план), проданных компании Гудзонова залива. Канадские рыси едят зайцев на снегоступах. В уравнениях Лотки-Вольтерра, также известный как уравнений хищник-жертва, являются парой первого порядка нелинейных дифференциальных уравнений, часто используется для описания динамики из биологических систем, в которых два вида взаимодействуют, один в качестве хищника, а другой как добычу. Популяции меняются со временем в соответствии с парой уравнений:
куда
Система уравнений Лотки – Вольтерра является примером модели Колмогорова, которая представляет собой более общую структуру, которая может моделировать динамику экологических систем с взаимодействиями хищник-жертва, конкуренцией, болезнями и взаимностью.
Хищник-жертва Лотки-Вольтерра модель была первоначально предложена Alfred J. Лотки в теории автокаталитических химических реакций в 1910 г. Это было эффективно логистическое уравнение, первоначально получаемые Ферхюльст. В 1920 году Лотка расширил модель через Андрея Колмогорова на «органические системы» на примере видов растений и травоядных животных, а в 1925 году он использовал уравнения для анализа взаимодействий хищник-жертва в своей книге по биоматематике. Та же самая система уравнений была опубликована в 1926 году Вито Вольтерра, математиком и физиком, заинтересовавшимся математической биологией. На расследование Вольтерра вдохновило его общение с морским биологом Умберто Д'Анкона, который в то время ухаживал за его дочерью, а позже стал его зятем. Д'Анкона изучил уловы рыбы в Адриатическом море и заметил, что процент пойманной хищной рыбы увеличился в годы Первой мировой войны (1914–18). Это озадачило его, так как за годы войны объемы рыбной ловли сильно сократились. Вольтерра разработал свою модель независимо от Лотки и использовал ее для объяснения наблюдения д'Анконы.
Позже модель была расширена, чтобы включить рост добычи в зависимости от плотности и функциональную реакцию в форме, разработанной К. С. Холлингом ; модель, которая стала известна как модель Розенцвейга – Макартура. Обе модели Лотка – Вольтерра и Розенцвейг – Макартур использовались для объяснения динамики естественных популяций хищников и добычи, таких как данные о рыси и зайцах-снегоступах компании Гудзонова залива и популяциях лосей и волков в Национальном парке острова Рояль.
В конце 1980-х годов появилась альтернатива модели Лотка – Вольтерра хищник – жертва (и ее обобщения, зависящие от общей жертвы), модель, зависимая от отношения, или модель Ардити – Гинзбурга. Обоснованность моделей, зависящих от добычи или соотношения, широко обсуждалась.
Уравнения Лотки – Вольтерра давно используются в экономической теории ; их первоначальная заявка обычно приписывается Ричарду Гудвину в 1965 или 1967 годах.
Модель Лотки – Вольтерры делает ряд предположений, не обязательно реализуемых по своей природе, об окружающей среде и эволюции популяций хищников и жертв:
В этом случае решение дифференциальных уравнений детерминировано и непрерывно. Это, в свою очередь, означает, что поколения хищника и жертвы постоянно пересекаются.
При умножении уравнение жертвы становится
Предполагается, что жертва имеет неограниченный запас пищи и размножается экспоненциально, если не подвергается нападению хищников; этот экспоненциальный рост представлен в приведенном выше уравнении членом αx. Предполагается, что скорость нападения хищников на добычу пропорциональна скорости, с которой встречаются хищники и жертва, это обозначено выше βxy. Если либо x, либо y равны нулю, хищничества быть не может.
С помощью этих двух членов приведенное выше уравнение можно интерпретировать следующим образом: скорость изменения популяции жертвы определяется ее собственной скоростью роста за вычетом скорости, с которой она становится жертвой.
Уравнение хищника становится
В этом уравнении δxy представляет рост популяции хищников. (Обратите внимание на сходство со скоростью хищничества; однако используется другая константа, поскольку скорость, с которой растет популяция хищников, не обязательно равна скорости, с которой они поедают добычу). Термин γy представляет собой скорость потери хищниками из-за естественной смерти или эмиграции, это приводит к экспоненциальному убыванию в отсутствие добычи.
Следовательно, уравнение выражает, что скорость изменения популяции хищника зависит от скорости, с которой он потребляет добычу, за вычетом его собственной смертности.
Уравнения имеют периодические решения и не имеют простого выражения в терминах обычных тригонометрических функций, хотя они вполне поддаются обработке.
Если ни один из неотрицательных параметров α, β, γ, δ не обращается в нуль, три могут быть включены в нормализацию переменных, чтобы оставить только один параметр: поскольку первое уравнение однородно по x, а второе по y, параметры β / α и δ / γ поглощаются нормализацией y и x соответственно, а γ - нормализацией t, так что только α / γ остается произвольным. Это единственный параметр, влияющий на характер решений.
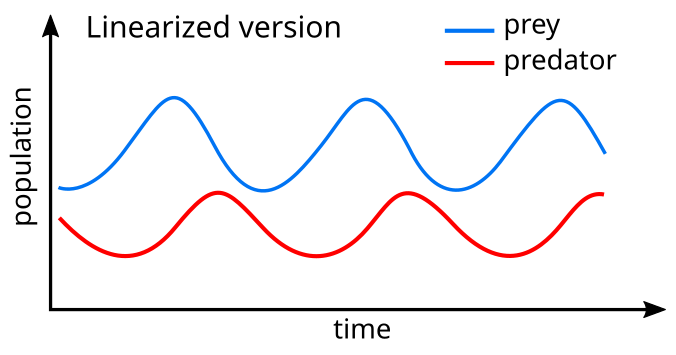
Линеаризация уравнений дает решение, аналогичное простому гармоническому движению с населением хищников задним, что хищный на 90 ° в цикле.
 Дополнительная информация: Предельный цикл
Дополнительная информация: Предельный цикл  Не говоря уже о динамике численности бабуинов и гепардов.
Не говоря уже о динамике численности бабуинов и гепардов.  График в фазовом пространстве для задачи хищник-жертва для различных начальных условий популяции хищников.
График в фазовом пространстве для задачи хищник-жертва для различных начальных условий популяции хищников. Предположим, есть два вида животных: бабуин (добыча) и гепард (хищник). Если начальные условия - 10 бабуинов и 10 гепардов, можно построить график эволюции двух видов с течением времени; учитывая параметры, согласно которым коэффициент роста и смертности павиана составляет 1,1 и 0,4, а у гепарда - 0,1 и 0,4 соответственно. Выбор временного интервала произвольный.
Можно также построить решения параметрически как орбиты в фазовом пространстве, не представляя время, но с одной осью, представляющей количество добычи, а другая ось, представляющая количество хищников за все время.
Это соответствует исключению времени из двух приведенных выше дифференциальных уравнений, чтобы получить одно дифференциальное уравнение
связывая переменные x и y. Решения этого уравнения представляют собой замкнутые кривые. Поддается разделению переменных : интегрирование
дает неявное отношение
где V - постоянная величина, зависящая от начальных условий и сохраняющаяся на каждой кривой.
Замечание: эти графики иллюстрируют серьезную потенциальную проблему с этим как с биологической моделью: для этого конкретного выбора параметров в каждом цикле популяция бабуинов сокращается до чрезвычайно низких чисел, но все же восстанавливается (в то время как популяция гепардов остается значительной на самом низком уровне. плотность павиана). Однако в реальных жизненных ситуациях случайные колебания дискретного числа особей, а также структуры семьи и жизненного цикла бабуинов могут привести к вымиранию бабуинов и, как следствие, гепардов. Эта проблема моделирования была названа «проблемой атто-лисицы», атто- лисица представляет собой условное 10 −18 лисицы.

Менее экстремальный пример охватывает:
α = 2/3, β = 4/3, γ = 1 = δ. Предположим, что x, y составляют тысячи каждый. Круги представляют собой начальные условия добычи и хищника от x = y = 0,9 до 1,8 с шагом 0,1. Фиксированная точка находится в точке (1, 1/2).
В модельной системе хищники процветают, когда добыча в изобилии, но в конечном итоге их запасы пищи опережают и сокращаются. Поскольку популяция хищников низка, популяция жертв снова увеличится. Эта динамика продолжается в демографическом цикле роста и спада.
Равновесие популяции происходит в модели, когда ни один из уровней популяции не меняется, т.е. когда обе производные равны 0:
Приведенная выше система уравнений дает два решения:
а также
Следовательно, есть два состояния равновесия.
Первое решение эффективно представляет исчезновение обоих видов. Если обе популяции равны 0, то они будут оставаться такими бесконечно. Второе решение представляет собой фиксированную точку, в которой обе популяции сохраняют свои текущие ненулевые числа и, в упрощенной модели, делают это бесконечно. Уровни населения, при которых достигается это равновесие, зависят от выбранных значений параметров α, β, γ и δ.
Устойчивость неподвижной точки в начале координат можно определить, выполнив линеаризацию с использованием частных производных.
Матрица Якоби модели хищник-жертва
и известна как матрица сообщества.
При оценке в установившемся состоянии (0, 0) матрица Якоби J становится
Собственные значения этой матрицы:
В модели α и γ всегда больше нуля, и поэтому знак собственных значений, указанных выше, всегда будет отличаться. Следовательно, неподвижная точка в начале координат является седловой точкой.
Важна стабильность этой неподвижной точки. Если бы он был стабильным, к нему могли бы быть привлечены ненулевые популяции, и поэтому динамика системы могла бы привести к исчезновению обоих видов во многих случаях начальных уровней популяции. Однако, поскольку фиксированная точка в начале координат является седловой и, следовательно, нестабильной, из этого следует, что вымирание обоих видов затруднено в модели. (Фактически, это могло произойти только в том случае, если жертва была полностью искоренена искусственно, в результате чего хищники умирали от голода. Если бы хищники были уничтожены, популяция жертвы неограниченно росла бы в этой простой модели.) Популяции жертвы и хищника могут стать бесконечно близким к нулю и все равно выздоравливать.
Вычисление J во второй фиксированной точке приводит к
Собственные значения этой матрицы:
Поскольку собственные значения являются чисто мнимыми и сопряженными друг другу, эта фиксированная точка должна быть либо центром замкнутых орбит в локальной окрестности, либо притягивающей или отталкивающей спиралью. В консервативных системах должны быть замкнутые орбиты в локальной окрестности неподвижных точек, которые существуют в минимумах и максимумах сохраняющейся величины. Выше было получено, что сохраняющаяся величина находится на орбитах. Таким образом, орбиты вокруг фиксированной точки являются замкнутыми и эллиптическими, поэтому решения являются периодическими, колеблющимися по небольшому эллипсу вокруг фиксированной точки с частотой и периодом.
Как показано в циркулирующих колебаниях на рисунке выше, кривые уровня представляют собой замкнутые орбиты, окружающие фиксированную точку: уровни популяций хищника и жертвы циклически изменяются и колеблются без затухания вокруг фиксированной точки с частотой.
Значение постоянной движения V, или, что то же самое, K = exp ( -V), может быть найдено для замкнутых орбит вблизи фиксированной точки.
Увеличение K перемещает замкнутую орбиту ближе к фиксированной точке. Наибольшее значение константы K получается при решении оптимизационной задачи
Таким образом, максимальное значение K достигается в стационарной (фиксированной) точке и составляет
где e - число Эйлера.

 и представляют собой мгновенные темпы роста двух популяций;
и представляют собой мгновенные темпы роста двух популяций;





















