
Войти
| Длиннопалая саламандра | |
|---|---|
 | |
| Статус сохранения | |
 Наименее опасны ( МСОП 3.1 ) Наименее опасны ( МСОП 3.1 ) | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Класс: | Амфибия |
| Порядок: | Уродела |
| Семья: | Амбистоматиды |
| Род: | Амбистома |
| Разновидность: | A. macrodactylum |
| Биномиальное имя | |
| Макродактилум амбистомы Бэрд, 1950 | |
| Подвиды | |
| Являюсь. columbianum A. m. croceum A. m. krausei A. m. macrodactylum A. m. сигиллатум | |
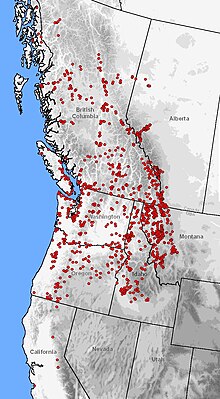 | |
| Распространение A. macrodactylum (красные точки) на западе Северной Америки | |
| Синонимы | |
| |
Длиннопалое саламандры ( Ambystoma macrodactylum) представляет собой мольное саламандры в семье Ambystomatidae. Этот вид, как правило, в зрелом возрасте составляет 4,1–8,9 см (1,6–3,5 дюйма) в длину, характеризуется пятнистой черной, коричневой и желтой пигментацией и длинным внешним четвертым пальцем на задних конечностях. Анализ ископаемых останков, генетика и биогеография позволяют предположить, что A. macrodactylum и A. laterale произошли от общего предка, который получил доступ к западным Кордильерам с потерей среднеконтинентального морского пути к палеоцену.
Распространение длиннопалой саламандры в основном на Тихоокеанском Северо-западе с высотным диапазоном до 2800 м (9 200 футов). Он обитает в различных средах обитания, в том числе в тропических лесах с умеренным климатом, хвойных лесах, прибрежных горных зонах, полынных равнинах, красных пихтовых лесах, полузасушливых полынных равнинах, степных равнинах и альпийских лугах вдоль скалистых берегов горных озер. Он живет в медленных ручьях, прудах и озерах во время фазы водного размножения. Длиннопалая саламандра впадает в спячку в холодные зимние месяцы, выживая за счет запасов энергии, хранящейся в коже и хвосте.
У этих пяти подвидов разная генетическая и экологическая история, фенотипически выраженная в различных цветах и типах кожи. Хотя длиннопалая саламандра классифицируются как разновидность наималейших опасений по МСОПУ, многие формы развития земли угрожают и отрицательно влияют на среду обитания саламандры в.
Ambystoma macrodactylum - представитель семейства Ambystomatidae, также известного как кротовые саламандры. Ambystomatidae произошли приблизительно 81 миллион лет назад (поздний мел ) от своего сестринского таксона Dicamptodontidae. Ambystomatidae также являются членами подотряда Salamandroida, который включает всех саламандр, способных к внутреннему оплодотворению. Сестринским видом A. macrodactylum является A. laterale, распространенный в восточной части Северной Америки. Однако филогенез Ambystomatidae на уровне видов является предварительным и требует дальнейшего тестирования.
Тело длиннопалого саламандры является смуглой черный с дорсальной полосой загар, желтой или оливково-зеленого цвета. Эта полоса также может быть разбита на несколько пятен. По бокам тела могут быть мелкие белые или бледно-голубые пятна. Брюшко темно-коричневого или черного цвета с белыми вкраплениями. Присутствуют корневые бугорки, но они не так развиты, как у других видов, таких как тигровая саламандра.
На яйца этого вида похожи на те, в связанной северо - западной саламандра ( А. грацильного) и тигровой саламандры ( A. tigrinum). Как и у многих земноводных, яйца длиннопалой саламандры окружены студенистой капсулой. Эта капсула прозрачная, благодаря чему эмбрион виден во время развития. В отличие от яиц A. gracile, здесь нет видимых признаков зеленых водорослей, что делает яичные желе зеленым цветом. В яйце зародыш длиннопалой саламандры темнее сверху и белее снизу по сравнению с эмбрионом тигровой саламандры, который от светло-коричневого до серого сверху и кремового цвета снизу. Яйца имеют диаметр около 2 мм (0,08 дюйма) или больше с широким внешним слоем желе. Перед вылуплением - как в яйце, так и в виде новорожденных личинок - у них есть балансиры - тонкие выступы кожи, выступающие по бокам и поддерживающие голову. В конечном итоге балансиры отваливаются, а их внешние жабры увеличиваются в размерах. Когда балансиры потеряны, личинки отличаются острым расширением жабр. Когда личинки созревают и превращаются в них, их конечности с пальцами становятся видимыми, а жабры рассасываются.
Кожа личинки пестрая с черной, коричневой и желтой пигментацией. Цвет кожи меняется по мере развития личинок, миграции пигментных клеток и их концентрации в разных частях тела. Пигментные клетки, называемые хроматофорами, происходят из нервного гребня. Три типа пигментных хроматофоров у саламандр включают желтые ксантофоры, черные меланофоры и серебристые иридиофоры (или гуанофоры). По мере созревания личинок меланофоры концентрируются вдоль тела и создают более темный фон. Желтые ксантофоры располагаются вдоль позвоночника и на верхней части конечностей. Остальная часть тела покрыта светоотражающими иридиофорами по бокам и снизу.
По мере того, как личинки метаморфизируются, они развивают пальцы из выступов зачатков конечностей. Полностью измененная длиннопалая саламандра имеет четыре пальца на передних конечностях и пять пальцев на задних конечностях. Его голова длиннее, чем ширина, а длинный внешний четвертый палец на задней конечности зрелых личинок и взрослых особей отличает этот вид от других, а также является этимологическим происхождением его специфического эпитета : макродактилум (греч. Makros = длинный и daktylos = палец).). Кожа взрослых особей имеет фон от темно-коричневого, темно-серого до черного с пятнами желтого, зеленого или тускло-красного цвета с точками и пятнами по бокам. Под конечностями, головой и телом саламандра белого, розоватого или коричневого цвета с более крупными белыми пятнами и более мелкими желтыми пятнами. Взрослые особи обычно имеют длину 3,8–7,6 см (1,5–3,0 дюйма).
Длиннопалая саламандра - это экологически разносторонний вид, обитающий в различных средах обитания, начиная от тропических лесов с умеренным климатом, хвойных лесов, горных прибрежных районов, полынных равнин, красных еловых лесов, полузасушливых полынных зарослей, степных равнин до альпийских лугов вдоль скалистых берегов гор. озера. Взрослые особи могут находиться в лесном подлеске, прячась под грубыми древесными обломками, камнями и в норах мелких млекопитающих. В весенний период размножения взрослых особей можно найти под обломками или на отмелях у рек, ручьев, озер и прудов. Часто бывают эфемерные воды.
Этот вид - одна из самых распространенных саламандр в Северной Америке, уступающая только тигровой саламандре. Его высота над уровнем моря составляет от уровня моря до 2800 метров (9 200 футов), охватывая широкий спектр растительных зон. Ареал включает изолированные эндемичные популяции в заливе Монтерей и Санта-Крус, Калифорния. Распространение воссоединяется в северо-восточной части Сьерра-Невады, непрерывно протянувшейся вдоль Тихоокеанского побережья до Джуно, Аляска, с популяциями, разбросанными по долинам рек Таку и Стикин. От побережья Тихого океана ареал простирается в продольном направлении до восточных предгорий Скалистых гор в Монтане и Альберте.
 Яичная масса длиннопалой саламандры с внешним слоем желе, соединяющим массу, и двумя внутренними капсулами, разделяющими каждое яйцо.
Яичная масса длиннопалой саламандры с внешним слоем желе, соединяющим массу, и двумя внутренними капсулами, разделяющими каждое яйцо. Как и у всех земноводных, жизнь длиннопалой саламандры начинается с яйца. На севере ареала яйца откладываются комковатыми массами вдоль травы, прутьев, камней или грязного субстрата спокойного пруда. Количество яиц в одной массе колеблется в размерах, возможно, до 110 яиц на одну группу. Самки вкладывают значительные средства в производство яиц, причем яичники составляют более 50% массы тела в предгнездовой период. У одной самки было обнаружено не более 264 яиц - большое количество, учитывая, что каждое яйцо имеет диаметр примерно 0,5 миллиметра (0,02 дюйма). Яичная масса удерживается вместе гелеобразным внешним слоем, защищающим внешнюю капсулу отдельных яиц. Яйца иногда откладывают поодиночке, особенно в более теплом климате к югу от границы с Канадой и США. Яичные желе вносят ежегодный запас биологического материала, который поддерживает химический состав и динамику питательных веществ в мелководных водных экосистемах и прилегающих лесных экосистемах. Яйца также являются средой обитания водяных форм, также известных как оомицеты.
 Являюсь. личинка macrodactylum с одноименными длинными четвертыми пальцами на задних лапах
Являюсь. личинка macrodactylum с одноименными длинными четвертыми пальцами на задних лапах Личинки вылупляются из оболочки яйца через две-шесть недель. Они прирожденные плотоядные животные, инстинктивно питающиеся мелкими беспозвоночными, которые движутся в их поле зрения. Пища включает мелких водных ракообразных ( кладоцеры, веслоногие рачки и остракоды ), водных двукрылых и головастиков. По мере развития они естественным образом питаются более крупной добычей. Чтобы увеличить свои шансы на выживание, некоторые особи отращивают большие головы, становятся каннибалами и питаются своими собственными товарищами по выводку.
После того, как личинки вырастут и созреют, по крайней мере, в течение одного сезона (личиночный период длится около четырех месяцев на побережье Тихого океана ), они поглощают свои жабры и превращаются в наземную молодь, которая бродит по лесному подлеску. О метаморфозе сообщалось еще в июле на уровне моря у A. m. croceum с октября по ноябрь и даже в январе. На более высоких высотах личинки могут перезимовать, развиваться и расти в течение дополнительного сезона перед метаморфозом. В озерах на больших высотах личинки могут достигать размеров 47 миллиметров (1,9 дюйма) от длины рыла до выходного отверстия (SVL) при метаморфозе, но на более низких высотах они развиваются быстрее и метаморфизируются, когда достигают 35-40 миллиметров (1,4–1,6 дюйма). SVL.
Во взрослом возрасте длиннопалые саламандры часто остаются незамеченными, потому что они ведут подземный образ жизни, копая, мигрируя и питаясь беспозвоночными в лесных почвах, гниющих бревнах, небольших норах грызунов или каменных трещинах. В рацион взрослого человека входят насекомые, головастики, черви, жуки и мелкая рыба. Саламандры охотился на на платочной змей, мелких млекопитающих, птиц и рыб. Взрослый особь может прожить 6–10 лет, при этом самые крупные особи весят примерно 7,5 грамма (0,26 унции), длина носа до вентиляционного отверстия достигает 8 см (3,1 дюйма), а общая длина достигает 14 см (5,5 дюйма).
История жизни длиннопалой саламандры сильно зависит от высоты и климата. Сезонные даты миграции в пруды-размножители и из них могут быть соотнесены с приступами продолжительных дождей, таяния льда или снега, достаточными для пополнения (часто) сезонных прудов. Яйца могут нереститься на низких высотах уже в середине февраля на юге Орегона, с начала января по июль на северо-западе Вашингтона, с января по март на юго-востоке Вашингтона и с середины апреля до начала мая в национальном парке Уотертон-Лейкс, Альберта. Сроки разведения могут сильно варьироваться; Следует отметить, что несколько яичных масс на ранних стадиях развития были обнаружены 8 июля 1999 г. вдоль границы провинции Британская Колумбия за пределами Джаспера, Альберта. Взрослые особи мигрируют сезонно, чтобы вернуться в свои родовые пруды, при этом самцы прибывают раньше и остаются дольше самок, а некоторые особи мигрируют по снежным берегам в теплые весенние дни. Гендерные различия (или половой диморфизм ) у этого вида проявляются только в период размножения, когда зрелые самцы демонстрируют увеличенную или выпуклую область вентиляции.
 Длиннопалые саламандры собираются во время сезона размножения под бревном непосредственно у берега пруда. Обратите внимание на диапазон оттенков кожи от серого до яркого.
Длиннопалые саламандры собираются во время сезона размножения под бревном непосредственно у берега пруда. Обратите внимание на диапазон оттенков кожи от серого до яркого. Время размножения зависит от высоты и широты места обитания саламандры. Обычно саламандры, живущие на более низких высотах, размножаются осенью, зимой и ранней весной. Высотные саламандры размножаются весной и в начале лета. Особенно в более высоких климатических условиях саламандры заходят в пруды и озера, в которых все еще плавает лед.
Взрослые особи собираются в больших количествах (gt; 20 особей) под камнями и бревнами вдоль непосредственной границы мест размножения и стремительно размножаются в течение нескольких дней. Подходящие места для размножения включают небольшие пруды, свободные от рыбы, болота, мелкие озера и другие стоячие водно-болотные угодья. Как и другие саламандры- амбистоматиды, они развили характерный танец ухаживания, когда они трутся телами и выделяют феромоны из подбородочной железы перед тем, как принять позу совокупления. Оказавшись в нужном положении, самец откладывает сперматофор, который представляет собой липкий стебель, на конце которого находится пакет со спермой, и направляет самку вперед для осеменения. Самцы могут спариваться более одного раза и могут откладывать до 15 сперматофоров в течение пятичасового периода. Брачный танец длиннопалой саламандры похож на другие виды Ambystoma и очень похож на A. jeffersonianum. У длиннопалой саламандры нет трения или ударов головой; самцы прямо подходят к самкам и хватаются за них, а самки стремятся быстро уплыть. Самцы обхватывают самку за передние конечности и трясут - это поведение называется амплексус. Самцы иногда во время размножения обнимают другие виды земноводных и тоже их трясут. Самец хватает только передними конечностями и никогда не использует задние конечности во время брачного танца, когда он трет подбородок из стороны в сторону, давя на голову самки. Женщина борется, но позже становится подавленной. Самцы увеличивают темп и движения, потирая ноздри самки, бока, а иногда и вентиляционное отверстие. Когда самка становится достаточно послушной, самец движется вперед, поднимая хвост над ее головой и взмахивая кончиком. Если самка принимает ухаживания самца, самец направляет ее мордочку в свою область вентиляции, в то время как оба движутся вперед жестко с волнообразными движениями таза. Когда самка следует, самец останавливается и откладывает сперматофор, а самка продвигается вперед вместе с самцом, чтобы поднять свой хвост и получить пакет со спермой. Полный танец ухаживания редко достигается с первой попытки. Самки откладывают яйца через несколько дней после спаривания.
 Длиннопалая саламандра с автоматическим хвостовым упором
Длиннопалая саламандра с автоматическим хвостовым упором  Длиннопалая саламандра с отметинами отрастания хвоста после потери
Длиннопалая саламандра с отметинами отрастания хвоста после потери В некоторых равнинных районах взрослые саламандры остаются активными всю зиму, за исключением холодов. Однако в холодные зимние месяцы в северных частях своего ареала длиннопалая саламандра зарывается ниже линии промерзания в грубый субстрат и впадает в спячку группами по 8–14 особей. Во время гибернации он выживает за счет энергетических запасов белка, которые хранятся в его коже и вдоль его хвоста. Эти белки выполняют второстепенную функцию как часть смеси или смеси кожных секретов, которые используются для защиты. При угрозе длиннопалая саламандра взмахивает хвостом и выделяет липкое белое молочно-белое вещество, которое является ядовитым и, вероятно, ядовитым. Цвет его кожи может служить предупреждением хищникам ( апосематизм ) о том, что он будет плохим на вкус. Цвет его кожи и узоры разнообразны: от темно-черного до красновато-коричневого фона, который испещрен пятнами или пятнами от бледно-красновато-коричневого, бледно-зеленого до ярко-желтой полосы. Взрослый также может уронить часть своего хвоста и ускользнуть, в то время как хвостовой укус действует как извивающаяся приманка; это называется аутотомией. Регенерации и восстановления роста хвоста один пример развития физиологии амфибий, что представляет большой интерес для медицинской профессии.
В то время как длиннопалая саламандра классифицируется МСОП как наименьшее беспокойство, многие формы освоения земель негативно влияют на среду обитания саламандры и ставят новые перспективы и приоритеты в ее охранную биологию. Приоритеты сохранения сосредоточены на уровне разнообразия популяций, которое сокращается в десять раз быстрее, чем исчезновение видов. Разнообразие на уровне населения - это то, что обеспечивает экосистемные услуги, такие как ключевую роль, которую саламандры играют в почвенных экосистемах, включая круговорот питательных веществ, который поддерживает водно-болотные и лесные экосистемы.
Две особенности жизненного цикла земноводных часто упоминаются как причина того, почему земноводные являются хорошими индикаторами состояния окружающей среды или «канарейками в угольной шахте». Как и все земноводные, у длиннопалой саламандры есть переход к водной и наземной жизни, а также полупроницаемая кожа. Поскольку они выполняют разные экологические функции в воде, чем на суше, потеря одного вида амфибий эквивалентна потере двух экологических видов. Второе понятие заключается в том, что земноводные, такие как длиннопалые саламандры, более восприимчивы к поглощению загрязняющих веществ, потому что они естественным образом поглощают воду и кислород через кожу. Однако обоснованность этой особой чувствительности к загрязнителям окружающей среды была поставлена под сомнение. Проблема более сложна, потому что не все земноводные одинаково восприимчивы к экологическому ущербу, потому что существует такой разнообразный набор историй жизни среди видов.
Популяции длиннопалых саламандр находятся под угрозой из-за фрагментации, интродуцированных видов и УФ-излучения. Лесное хозяйство, дороги и другие земельные участки изменили среду, в которую мигрируют земноводные, и увеличили смертность. В таких местах, как Национальный парк Уотертон-Лейкс, проложен подземный переход в автомобильном туннеле, чтобы обеспечить безопасный проход и поддержать миграционную экологию этого вида. [2] Распространение длиннопалой саламандры во многом совпадает с лесной промышленностью, доминирующим ресурсом, поддерживающим экономику Британской Колумбии и западной части Соединенных Штатов. Длиннопалые саламандры изменят миграционное поведение, и на них негативно повлияют методы ведения лесного хозяйства, не обеспечивающие значительных буферов управления и защиты для небольших водно-болотных угодий, где размножаются саламандры. Население вблизи долины реки Мис, Альберта, было потеряно из-за расчистки и осушения водно-болотных угодий для сельского хозяйства. Форель, завезенная для спортивного рыболовства в некогда безрыбные озера, также уничтожает популяции длиннопалых саламандр. Интродуцированная золотая рыбка питается икрой и личинками длиннопалых саламандр. Повышенное воздействие УФ-В излучения является еще одним фактором, влияющим на глобальное сокращение численности земноводных, и длиннопалые саламандры также подвержены этой угрозе, что увеличивает частоту уродств и снижает их выживаемость и темпы роста.
Подвид Ambystoma macrodactylum croceum ( длиннопалая саламандра Санта-Крус ) вызывает особую озабоченность, и в 1967 году ему была предоставлена защита в соответствии с Законом США об исчезающих видах. Этот подвид обитает в узком диапазоне ареалов в округах Санта-Крус и Монтерей, штат Калифорния. До получения защиты некоторые оставшиеся популяции находились под угрозой развития. Подвид является экологически уникальным, с уникальным и неправильным рисунком кожи на спине, уникальной устойчивостью к влаге, а также является эндемиком, географически изолированным от остального ареала вида. Другие подвиды включают A. m. columbianum, A. m. krausei, A. m. macrodactylum и A. m. сигиллатум.
Предки этого вида происходят из восточной части Северной Америки, где видовое богатство амбистоматид наиболее высоко. Следующая биогеографическая интерпретация происхождения A. macrodactylum в западной части Северной Америки основана на описании окаменелостей, генетики и биогеографии. Ближайшим из ныне живущих родственных видов длиннопалой саламандры является A. laterale, произрастающий на северо-востоке Северной Америки. Ambystomatidae были изолированы к юго-востоку от Срединно -континентального или Западного внутреннего морского пути в меловом периоде (~ 145,5–66 млн лет назад). В то время как три других вида Ambystomatidae ( A. tigrinum, A. californiense и A. gracile ) имеют перекрывающиеся ареалы в западной части Северной Америки, ближайшим родственным видом длиннопалой саламандры является A. laterale, уроженец северо-востока Северной Америки. Было высказано предположение, что A. macrodactylum произошел от A. laterale после палеоцена (~ 66–55,8 млн лет) с потерей Западного внутреннего морского пути, открывшего путь для общего предка в Западные Кордильеры. Когда-то обитавшие в горных районах западной части Северной Америки, видам приходилось сталкиваться с динамичной пространственной и композиционной экологией, реагирующей на изменения высоты по мере роста гор и изменения климата. Например, Pacific Northwest стал прохладнее палеоцена, проложив путь для умеренного леса, чтобы заменить теплый тропический лес из мела. Сценарий отделения A. macrodacylum и других западных видов умеренного пояса от их восточных собратьев предполагает поднятие Скалистых гор в позднем олигоцене до миоцена. Орогения создал климатический барьер путем удаления влаги из потока воздуха и западного сушат в midcontinental зону, из южной Альберты в Мексиканском заливе.
Предки современных саламандр, вероятно, смогли рассредоточиться и мигрировать в места обитания Скалистых гор и прилегающих территорий к эоцену. Мезические леса образовались в западной части Северной Америки к середине эоцена и достигли своего современного распространения к началу плиоцена. Умеренные лесные долины и горная среда этих периодов времени (от палеогена до неогена ) обеспечили бы физико-географические и экологические особенности, поддерживающие аналоги современных местообитаний Ambystoma macrodactylum. Каскад Диапазон вырос в течение середины плиоцена и создал дождь теневого эффекта, вызывающий xerification из Колумбии бассейна, а также изменил диапазоны умеренных мезонных экосистем на возвышенностях. Возникновение Каскадов, вызвавшее ксерификацию бассейна Колумбии, является важной биогеографической особенностью западной части Северной Америки, которая разделила многие виды, включая A. macrodactylum, на прибрежные и внутренние линии.
Существует пять подвидов длиннопалой саламандры. Подвиды различаются по их географическому положению и рисункам на спинной полосе; Дензел Фергюсон дает биогеографический отчет о структуре и морфологии кожи; на основе этого анализа он представил два новых подвида: A. macrodactylum columbianum и A. m. сигиллатум. Ареалы подвидов проиллюстрированы в путеводителях Роберта Стеббина по амфибиям.
 Ареал A. macrodactylum и ареалы распространения подвидов.
Ареал A. macrodactylum и ареалы распространения подвидов.  Слева: на спинной нижней стороне под его мордой находятся подбородочные железы, которые выделяют гормоны, когда самцы трутся о самку. Они расположены под белыми пятнами. Справа: область мужского отверстия (выводное и репродуктивное отверстие) имеет увеличенную анатомию и увеличенные сосочки (на фото). Женское отверстие не такое большое и часто более гладкое, без складок сосочков.
Слева: на спинной нижней стороне под его мордой находятся подбородочные железы, которые выделяют гормоны, когда самцы трутся о самку. Они расположены под белыми пятнами. Справа: область мужского отверстия (выводное и репродуктивное отверстие) имеет увеличенную анатомию и увеличенные сосочки (на фото). Женское отверстие не такое большое и часто более гладкое, без складок сосочков. Сводка отличительных рисунков кожи и морфологических особенностей подвидов включает:
Анализ митохондриальной ДНК выявляет несколько разные диапазоны для подвидов клонов. Генетический анализ, например, выявляет дополнительную картину глубокой дивергенции в восточной части ареала. Пространственное распределение популяций и генетика этого вида связаны пространственно и исторически через взаимосвязанные системы горных и умеренных долин на западе Северной Америки. Верность размножения длиннопалых саламандр ( филопатрия ) и другие виды миграционного поведения снижают скорость расселения между регионами, например, в пределах горных бассейнов. Этот аспект их поведения ограничивает поток генов и увеличивает степень и скорость генетической дифференциации. Генетическая дифференциация между регионами у длиннопалых саламандр выше, чем у большинства других групп позвоночных. Естественные перерывы в диапазоне расселения и миграции происходят там, где экосистемы переходят в более сухие ксерические низменности (например, климат прерий ) и на замерзшей или суровой местности на больших высотах (2200 метров (7200 футов)).
Томпсон и Рассел обнаружили еще одно эволюционное происхождение, которое берет свое начало в ограниченном ледниками районе гор Салмон-Ривер, штат Айдахо. С приходом голоценового межледниковья, примерно 10 000 лет назад, плейстоценовые ледники отступили и открыли миграционный путь, связывающий эти южные популяции с северными районами, где они в настоящее время пересекаются с A. m. krausei и мигрировали на север в долину реки Мис (Канада). Фергюсон также отметил взаимную градацию в той же географической области, но между морфологическими подвидами A. m. columbianum и A. m. krausei, которые проходят параллельно хребтам Биттеррут и Селкирк. Томпсон и Рассел предполагают, что эта зона контакта находится между двумя разными подвидами линий, поскольку A. m. columbianum географически изолирована и ограничена центральными горами Орегона.
![]() Данные, относящиеся к Ambystoma macrodactylum на Wikispecies
Данные, относящиеся к Ambystoma macrodactylum на Wikispecies ![]() Media, связанные с Ambystoma macrodactylum на Викискладе?
Media, связанные с Ambystoma macrodactylum на Викискладе?