
Войти
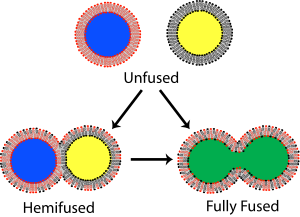 Иллюстрация слияния липидных пузырьков, показывая два возможных исхода: гемифузия и полное слияние. При гемифузии смешиваются только наружные двухслойные листочки. При полном слиянии обе створки и внутреннее содержимое смешиваются.
Иллюстрация слияния липидных пузырьков, показывая два возможных исхода: гемифузия и полное слияние. При гемифузии смешиваются только наружные двухслойные листочки. При полном слиянии обе створки и внутреннее содержимое смешиваются. В мембранной биологии, слияние - это процесс, посредством которого два изначально различных липидных бислоя сливаются их гидрофобные ядра, в результате чего получается одна взаимосвязанная структура. Если это слияние полностью проходит через обе створки обоих бислоев, образуется водный мостик, и внутреннее содержимое двух структур может смешиваться. Альтернативно, если только одна листочка из каждого бислоя участвует в процессе слияния, говорят, что бислои гемифузированы. При гемифузии липидные составляющие наружной створки двух бислоев могут смешиваться, но внутренние створки остаются отдельными. Водное содержимое, заключенное в каждом бислое, также остается разделенным.
Слияние участвует во многих клеточных процессах, особенно у эукариот, поскольку эукариотическая клетка широко разделена липидными двухслойными мембранами. Экзоцитоз, оплодотворение яйцеклетки спермой и перенос продуктов жизнедеятельности в лизосомы - вот лишь некоторые из многие эукариотические процессы, основанные на той или иной форме слияния. Слияние также является важным механизмом транспорта липидов от места их синтеза к мембране, где они необходимы. Даже проникновение патогенов может регулироваться слиянием, поскольку многие вирусы с двухслойным покрытием имеют специальные слитые белки для проникновения в клетку-хозяина.
В процессе слияния есть четыре основных шага, хотя каждый из этих шагов фактически представляет собой сложную последовательность событий. Во-первых, задействованные мембраны должны агрегироваться, приближаясь друг к другу с точностью до нескольких нанометров. Во-вторых, два бислоя должны войти в очень тесный контакт (в пределах нескольких ангстрем). Чтобы достичь этого тесного контакта, две поверхности должны стать, по крайней мере, частично обезвоженными, так как обычно присутствующая связанная поверхностная вода заставляет двухслойные слои сильно отталкиваться на таком расстоянии. В-третьих, дестабилизация должна развиваться в одной точке между двумя бислоями, вызывая сильно локализованную перестройку двух бислоев. Наконец, по мере роста этого точечного дефекта компоненты двух бислоев смешиваются и диффундируют от места контакта. В зависимости от того, происходит ли гемифузия или полное слияние, внутреннее содержимое мембран также может смешиваться в этот момент.
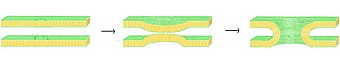 Схематическая иллюстрация процесса слияния посредством образования стебля.
Схематическая иллюстрация процесса слияния посредством образования стебля. Точные механизмы, лежащие в основе этой сложной последовательности событий, до сих пор остаются предметом споров. Чтобы упростить систему и позволить более точное исследование, многие эксперименты были выполнены in vitro с синтетическими липидными везикулами. Эти исследования показали, что двухвалентные катионы играют решающую роль в процессе слияния, связываясь с отрицательно заряженными липидами, такими как фосфатидилсерин, фосфатидилглицерин и кардиолипин. Одна из ролей этих ионов в процессе слияния - экранировать отрицательный заряд на поверхности бислоя, уменьшая электростатическое отталкивание и позволяя мембранам приближаться друг к другу. Однако это явно не единственная роль, поскольку существует широко задокументированная разница в способности Mg по сравнению с Ca вызывать слияние. Хотя Mg вызывает обширную агрегацию, он не вызывает слияния, в то время как Ca вызывает и то, и другое. Было высказано предположение, что это несоответствие связано с разницей в степени обезвоживания. Согласно этой теории, ионы кальция сильнее связываются с заряженными липидами, но менее сильно с водой. Возникающее в результате вытеснение кальция водой дестабилизирует поверхность раздела липид-вода и способствует тесному межслойному контакту. Недавно предложенная альтернативная гипотеза состоит в том, что связывание кальция вызывает дестабилизирующее боковое напряжение. Каким бы ни был механизм индуцированного кальцием слияния, начальное взаимодействие явно электростатическое, поскольку цвиттерионные липиды не восприимчивы к этому эффекту.
В процессе слияния головная липидная группа - это не только участвует в плотности заряда, но может влиять на дегидратацию и зарождение дефектов. Эти эффекты не зависят от воздействия ионов. Наличие незаряженного фосфатидилэтаноламина (PE) головной группы увеличивает слияние при включении в бислой фосфатидилхолина. Некоторые объясняют это явление эффектом обезвоживания, аналогичным влиянию кальция. Головная группа из полиэтилена связывает воду менее плотно, чем поликарбонат, и поэтому может облегчить закрытие соединения. Альтернативное объяснение состоит в том, что физическая, а не химическая природа PE может способствовать слиянию. Согласно гипотезе стебля слияния, сильно изогнутый мост должен образоваться между двумя бислоями, чтобы слияние произошло. Поскольку PE имеет небольшую головную группу и легко образует инвертированные фазы мицелл, он, согласно модели стебля, должен способствовать образованию этих стеблей. Дополнительным свидетельством, приведенным в пользу этой теории, является тот факт, что было показано, что определенные липидные смеси поддерживают слияние только при повышении температуры выше температуры перехода этих инвертированных фаз. Эта тема также остается спорной, и даже если существует криволинейная структура присутствует в процессе сварки, есть дебаты в литературе более, является ли она кубической, гексагональной или более экзотической расширенной фазой.
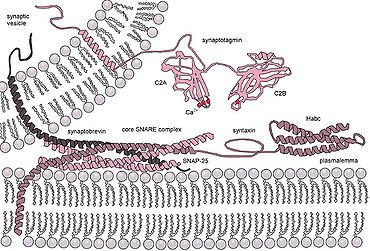 Схема действия белков SNARE, стыкующих везикулу для экзоцитоза. Дополнительные версии белка на везикуле и целевой мембране связываются и обвиваются вокруг друг друга, сближая в процессе два бислоя.
Схема действия белков SNARE, стыкующих везикулу для экзоцитоза. Дополнительные версии белка на везикуле и целевой мембране связываются и обвиваются вокруг друг друга, сближая в процессе два бислоя. Ситуация еще более усложняется при рассмотрении слияния in vivo, поскольку биологическое слияние почти всегда регулируется действие мембраносвязанных белков. Первыми из этих белков, которые были изучены, были слитые белки вируса, которые позволяют заключенному в оболочку вирусу вставлять свой генетический материал в клетку-хозяина (оболочечные вирусы - это вирусы, окруженные липидным бислоем; некоторые другие имеют только белковая оболочка). В целом, существует два класса слитых белков вирусов: кислотные и pH-независимые. pH независимые гибридные белки могут функционировать в нейтральных условиях и сливаться с плазматической мембраной, позволяя вирусам проникать в ячейка. Вирусы, использующие эту схему, включали ВИЧ, кори и герпес. Кислотные слитые белки, такие как те, что обнаружены в гриппе, активируются только при низком pH кислых эндосом и должны сначала быть эндоцитозированы, чтобы проникнуть в клетку.
Эукариотические клетки используют совершенно разные классы слитых белков, наиболее изученными из которых являются SNARE. Белки SNARE используются для направления всего везикулярного внутриклеточного транспорта. Несмотря на годы исследований, многое еще неизвестно о функции этого класса белков. Фактически, до сих пор ведутся активные дебаты относительно того, связаны ли SNAREs с ранней стыковкой или участвуют позже в процессе слияния, облегчая гемифузию. Даже после того, как будет освещена роль SNARE или других специфических белков, единое понимание слитых белков маловероятно, поскольку внутри этих классов существует огромное разнообразие структур и функций, а сохранено очень мало тем.
В исследованиях молекулярной и клеточной биологии часто бывает желательно искусственно вызвать слияние. Хотя это может быть достигнуто с добавлением кальция, как обсуждалось ранее, эта процедура часто невозможна, потому что кальций регулирует многие другие биохимические процессы, и его добавление может вызвать серьезные затруднения. Кроме того, как уже упоминалось, кальций вызывает массивную агрегацию, а также слияние. Добавление полиэтиленгликоля (PEG) вызывает слияние без значительной агрегации или биохимического нарушения. Эта процедура теперь широко используется, например, путем слияния B-клеток с миеломными клетками. Полученная «гибридома » из этой комбинации экспрессирует желаемое антитело, как определено участвующими В-клетками, но бессмертна из-за миеломного компонента. Механизм слияния PEG окончательно не идентифицирован, но некоторые исследователи полагают, что PEG, связывая большое количество молекул воды, эффективно снижает химическую активность воды и, таким образом, обезвоживает головные группы липидов. Слияние также может быть вызвано искусственно посредством электропорации в процессе, известном как электрослияние. Считается, что это явление является результатом энергетически активных краев, образованных во время электропорации, которые могут действовать как локальные дефектные точки для зарождения стеблей между двумя бислоями.
Существует два уровня слияния: смешивание мембранных липидов и смешивание содержимого. Анализы слияния мембран сообщают либо о смешивании мембранных липидов, либо о смешивании водного содержимого слитых объектов.
Анализы, оценивающие смешение липидов, используют эффекты, зависящие от концентрации, такие как безызлучательный перенос энергии, тушение флуоресценции и образование эксимера пирена.
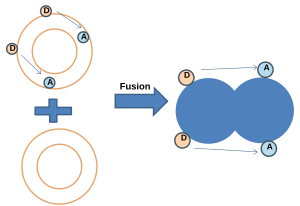 (1.) Иллюстрация анализа смешения липидов на основе резонансного переноса энергии Фёрстера.
(1.) Иллюстрация анализа смешения липидов на основе резонансного переноса энергии Фёрстера.  (3.) Иллюстрация анализа смешения липидов, основанного на самогашении флуоресценции.
(3.) Иллюстрация анализа смешения липидов, основанного на самогашении флуоресценции. Смешивание водного содержимого из везикул в результате лизиса, слияния или физиологической проницаемости может быть обнаружено флуорометрическим методом с использованием растворимых индикаторов с низкой молекулярной массой.
 (1.) Иллюстрация анализа смешивания содержимого на основе пары тушения флуоресценции ANTS / DPX.
(1.) Иллюстрация анализа смешивания содержимого на основе пары тушения флуоресценции ANTS / DPX.  (2.) Иллюстрация анализа смешивания содержимого на основе пары усиления флуоресценции Tb / DPA.
(2.) Иллюстрация анализа смешивания содержимого на основе пары усиления флуоресценции Tb / DPA.